
A közegészségügy rendezését célzó 1876. évi XIV. törvénycikk egyik legvitatott része a “védhimlőoltásokról” rendelkező XIII. fejezet volt, amely kötelezővé tette az addig csak opcionális himlőoltásokat. A pontos megfogalmazás szerint:
93. § A védhimlőoltás, esetleg a felnőttek ujra-oltása, évenként minden községben teljesitendő. Szülők és gyámok, átalában mindazok, kik gyermekekről gondoskodni tartoznak, kötelesek ezeket éltüknek első évében, a mennyiben valóságos himlőt ki nem állottak volna, beoltani. Mennyiben halasztható el egyesek beoltása, az az orvos megitélésétől függ. […]
95. § A himlőoltás teljesitése s a himlőanyag szedése és megőrzése körül szükséges elővigyázat 1-10 forintig, esetleg 2 napi fogságig terjedhető büntetés terhe alatt pontosan megtartandó.
A törvény parlamenti vitájában az oltások kötelező volta vonta maga után a legtöbb kritikát; a vitáról tudósító Pesti Napló beszámolójában:
A törvényjavaslat 93-ik szakaszához Hammersberg [Jenő] tudvalevőleg azon módositványt nyujtá be, hogy a himlőoltás kötelezővé ne tétessék. Vidlicskay József pártolja Hammersberg módositványát, a mennyiben a törvényjavaslat szövege által az egyéni szabadságot látja korlátolva. Knöpfler Vilmos a törvényjavaslat szövegét pártolja. Elismeri ugyan, hogy a tudomány még nem állapodott meg végleg a himlőoltás physiologiai hatásáról; de a gyógygyakorlat határozottan annak czélszerüsége mellett szól. Utal Angliára, hol a himlőoltás szintén kötelezőleg van behozva s elfogadtatni kívánja a törvényjavaslat rendelkezését. Tisza László az eredeti szöveg mellett szól főleg a ragály meggátlása szempontjából. A ház az előadó felszólalása után elfogadja az eredeti szöveget.
A szigor oka nem volt véletlen: a himlő még a korszak egyik leggyilkosabb betegsége volt, ma már elképzelhetetlen mértékű terhet róva a családokra, hiszen a rendszeresen ismétlődő himlőjárványok mellett szinte kikerülhetetlen volt, hogy valaki el ne kapja a betegséget. S mivel a betegség mortalitása közel 30%-os volt, különösen a 15 év alatti gyerekeket érintve, nem lehetett egyetlen legyintéssel elintézni az ebből származó társadalmi terheket, különösen, hogy ekkor már Edward Jenner munkásságának köszönhetően közel 80 éve létezett egy hatékony védekezés védőoltás formájában. (Megj: talán mindenkit segít orientálni abban, hogy ez a ma kvázi elfeledett betegség egykor mennyire pusztító volt, hogy az Our World in Data vonatkozó összefoglalása szerint létezésének utolsó 100 éve során 500 millió (!!) haláleset volt a himlő számlájára írható.)
A tervezett szigor azonban nem váltotta be a hozzá függő reményeket, jelentős passzív ellenállás mutatkozott, így nem véletlen, hogy aztán az 1887. évi XII. törvénycikk még szigorúbb szabályozásokkal igyekezett valóban, kikerülhetetlenül kötelezővé tenni az oltásokat – például a közösségbe járást oltáshoz, vagy öt évente történő újraoltáshoz kötötték. Hogy kevésbé lehessen ellógni és kikerülni az eljárást, a belügyminiszter körrendeletet hozott, amely értelmében az oltásokat egy bizottság előtt kellett beadni.
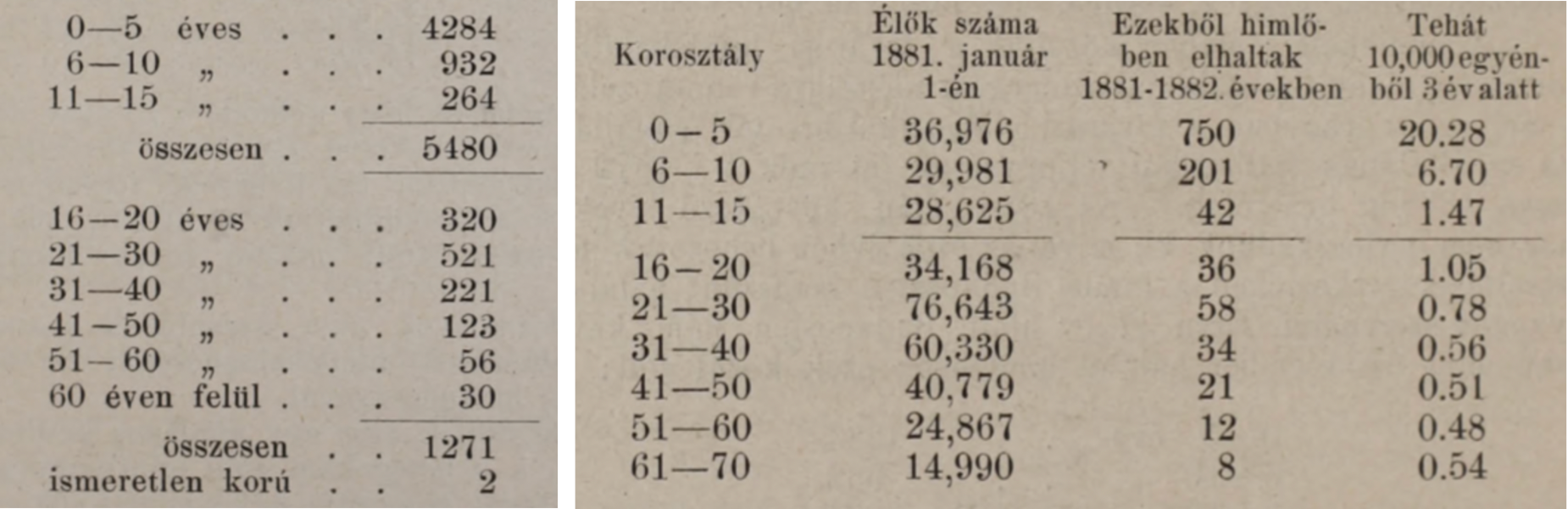
Ennek megfelelően tulajdonképpen az 1888-as év oltáskampánya tekinthető annak a pontnak, amikor Magyarországon is valóban kötelező és teljeskörű lett az himlőellenes védoltás. Aminek az eredményei meglepően hamar látszottak is. Amint azt Kőrösy József 1897-es “A himlődoltás véderejéről” szóló művében nagyon szemléletesen feldolgozta, mindazon országokban ahol bevezették a kötelező oltást, a “himlőhalálozás” töredékére hullott vissza, még a járványos években is. Az európai éllovas, Svédország példája különösen látványos – és ma már azt is tudjuk, hogy 1895-re, a kontinens egyik (ha nem az) első országa lettek, ahol az évezredek óta vissza-visszatérő betegség megszűnt endemikusnak lenni, vagyis csak akkor jelentett problémát, ha valahonnan behurcolták.
Nem mindenkit sikerült így sem megmenteni, de azért nem túlzás azt állítani, hogy néhány évtized alatt milliós számban éltek azok az emberek, akik a himlőoltás nélkül nem élték volna meg a felnőttkort. S mivel, úgy tűnik, a Monarchián belül ekkor még Budapest működött Bécs “laboratóriumaként”, pont az osztrák fővárossal összevetve is jól látható, hogy a magyar fővárosban már szűk évtized alatt is gyakorlatilag megszűnt a himlő, mint közegészségügyi probléma.
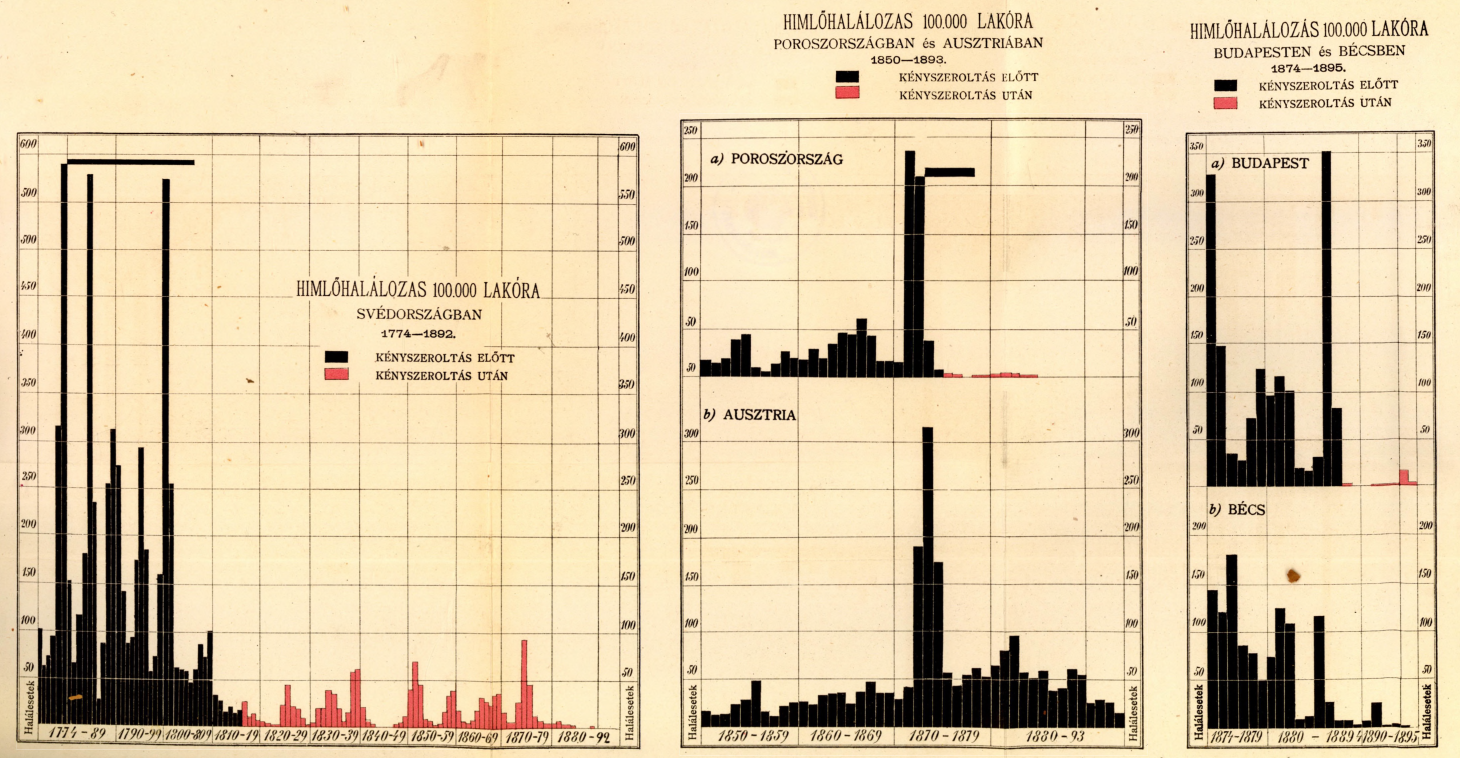
Kőrösy korának egyik legjelesebb statisztikusa volt, a Fővárosi Statisztikai Hivatal első vezetője, akinek adatsorai és indexszámítási eljárásai nemzetközi hírnevet hoztak és például magára Francis Galtonra is nagy hatással voltak. Említett műve a Jenner-féle védőoltás százéves évfordulójára készült el és rövidsége, illetve könnyen követhető nyelvezete miatt még egész biztosan ma is bátran ajánlható mindenki számára, akit a járványtan, vagy a közegészségügy érdekel.
Külön említést érdemel a harmadik fejezet („Az oltóellenes iskola statisztikai bizonyítékainak dialektikája”), amely számos tudománykommunikátor örök (és ma különösen érvényes) dilemmáját fogalmazza meg, azaz egy igazi kutató nem söpörheti le az egyes állításokat és “leleplezéseket”, ugyanakkor ezek tételes cáfolata aránytalanul nagy energiát igényel, főleg, hogy kétséges, mennyire győzhetőek meg a szkeptikusok:
Ha az oltásellenes nézeteket nem elfogultan fogadjuk, nem viseltetünk már kezdettől fogva gyanúval az illető szerzők iránt; ha általában hajlandók vagyunk oly nézetek lehetőségét és igazoltságát is elismerni, a melyek saját felfogásainkkal, esetleg tudományos meggyőződésünkkel is egyenes ellentétben állanak: kell hogy valamely komolyabb antivaccinatorikus munka átolvasása után, bizonyos nyugtalanság vegyen rajtunk erőt és hogy a himlőoltás hatásos voltában való erős bizalmunkat némileg megingatva érezzük. Fokozza ezen nyugtalanságot a használt bizonyítékok sokfélesége, a legkülönbözőbb, gyakran mesterkélt és emellett kaleidoskopszerűleg változó érvelések tarkasága. […]
Tisztességes, őszinte polémiánál azon kell lennünk, hogy magunkat az ellenfél gondolatmenetébe beletaláljuk. Hacsak enmagunkat csalhatatlanoknak nem tartjuk, ha nem hiszsziik, hogy az egyszer felállított tudományos állítások többé meg nem dönthetők, ugyan hogyan ne éreznék az oltásba vetett bizodalmunkat megingatva, ha pl. látjuk, hogy egy nagy kórháznak igazgatója és főorvosa — hivatkozással az ő saját tapasztalataira, sőt azoknak idézése mellett! — a himlőoltás meddőségét vallja […].
A mi engem illet, sok időn át kerültem az ezen vitába való elegyedést, kényelmesebbnek találván az orvosi tudomány vezérférfiainak véleményében egyszerűen belényugodni. Midőn azonban a hetvenes években, a fővárosban is kitört nagy himlőjárvány alatt, abbeli szándékom, hogy családom tagjait ez alkalommal újból oltassam, az eziránt megkeresett több orvosbarátom félreismerhetlen kételyével találkozott, úgy a tudományos mint az egyéni érdekeltség arra indított, hogy ezen sokat vitatott kérdésnek statisztikai alapját mégis megvizsgáljam.
Kőrösy azonban ezt követően azt gyönyörűen bemutatja azt is, hogy az oltásellenesek látszólagos statisztikai bizonyítékok mögött milyen csúsztatások vannak: például úgy dobálóznak halálesetek abszolút számaival, hogy közben nem vesznek tudomást egy-egy város robbanásszerű növekedéséről (amely miatt, ha 100 000 lakosra számítanák a halálozást máris nem működne az érvrendszerük), vagy egy-egy oltatlan terület járványmentes időszakának adatait vetik össze járványsújtott, de beoltott területekével. Ha viszont ezeket mind korrigáljuk, akkor „megbízható statisztika mellett éppenséggel lehetetlen az oltás véderejét megczáfolni.”
Ilyen és hasonló logikai bukfencek és csúsztatások, almák és körték összehasonlítása természetesen mindenki számára ismerős lesz, aki a napi híreket követi. Ha mondjuk egy sikeresen védekező országban az egymillió főre eső (összesen) 10 haláleset 100-ra fut fel, az már tízszeres növekedést jelent, amihez képest az egymillió főre eső 3100 és 3300 közti különbség szinte elhanyagolhatónak tűnik, pedig abszolút értékben lényegesen több halálesetet jelent. Vagy hasonló módon, egy nagyon nagy átoltottságú országban, a tünetes koronavírusos betegek jelentős, akár az oltatlanokéval összemérhető része lesz oltott – csak hát nem mindegy mekkora arányokról beszélünk. Ismét csak Kőrösy megfogalmazásában:
A leggyakoribb hibaforrást képezi az észlelt tömeg absolut nagyságának megnemfigyelése. A statisztikának a különböző nagyságú tömegeket mind egyforma nevezőre kell hoznia: ez okból tulajdonképen mindig csak arányszámokban, mindig csak százalékokban gondolkozik.
Az egy és negyed százada írt kis pamflet ma is érvényes sorainak olvasása közben két gondolat fogalmazódhat meg bennünk: egyrészt mennyire tragikus, hogy ugyanazokat a köröket futjuk ismét és ismét le, másrészt, konkrétan a himlő esetében hihetetlenül hatékony volt a kötelező oltásokra alapozó nemzetközi kampány és ennek megfelelően sokkal, de sokkal halkabbak lettek konkrétan a himlőoltás hatékonyságát és fontosságát megkérdőjelező hangok. Talán ez a legtöbb, amiben a koronavírusos oltások esetében is bízhatunk.
(A témában lásd még korábbi posztjainkat az oltások hatékonyságáról [1, 2, 3], az oltás vs. szabadságjogok kérdéskörről, illetve az oltáskommunikáción nehézségeiről [1, 2]).



 Ha Darwinról és hozzá közel álló madarakról beszélünk, akkor talán nem is feltétlenül a kötelező újggyakorlatként felemlegetett galapagosi pintyeket (aka. “Darwin pintyek”)
Ha Darwinról és hozzá közel álló madarakról beszélünk, akkor talán nem is feltétlenül a kötelező újggyakorlatként felemlegetett galapagosi pintyeket (aka. “Darwin pintyek”) 



;_Fragment_of_a_Letter_or_Document_(verso)_MET_DP800417.jpg){kind=link}
{kind=link}