 Bár a Fajok eredete igazából nem az ember eredetéről szól, mégis mindenki tudta és érezte már a megjelenésekor, hogy ez az az igazi kérdés, amit a benne leírt tudományos elmélet feszeget. Darwin, a rá jellemző visszafogottsággal, ebben a könyvében csak egy rövidke utalást tesz a kérdésre (“Fény derül majd az ember eredetére és történetére. (Light will be thrown on the origin of man and his history)”, amolyan beharangozóként későbbi, másik főművéhez, ami Az ember származása és az ivari kiválás lenne. Mindez jól tükrözi, hogy már bő másfél évszázada (ha pont nem “már a görögök óta”) az egyik legfontosabb kérdés, ami sok biológiai kutatás középpontjában is áll, arra keresi a választ, hogy mi az, ami minket fajként különlegessé tesz és megkülönböztet más fajoktól.
Bár a Fajok eredete igazából nem az ember eredetéről szól, mégis mindenki tudta és érezte már a megjelenésekor, hogy ez az az igazi kérdés, amit a benne leírt tudományos elmélet feszeget. Darwin, a rá jellemző visszafogottsággal, ebben a könyvében csak egy rövidke utalást tesz a kérdésre (“Fény derül majd az ember eredetére és történetére. (Light will be thrown on the origin of man and his history)”, amolyan beharangozóként későbbi, másik főművéhez, ami Az ember származása és az ivari kiválás lenne. Mindez jól tükrözi, hogy már bő másfél évszázada (ha pont nem “már a görögök óta”) az egyik legfontosabb kérdés, ami sok biológiai kutatás középpontjában is áll, arra keresi a választ, hogy mi az, ami minket fajként különlegessé tesz és megkülönböztet más fajoktól.
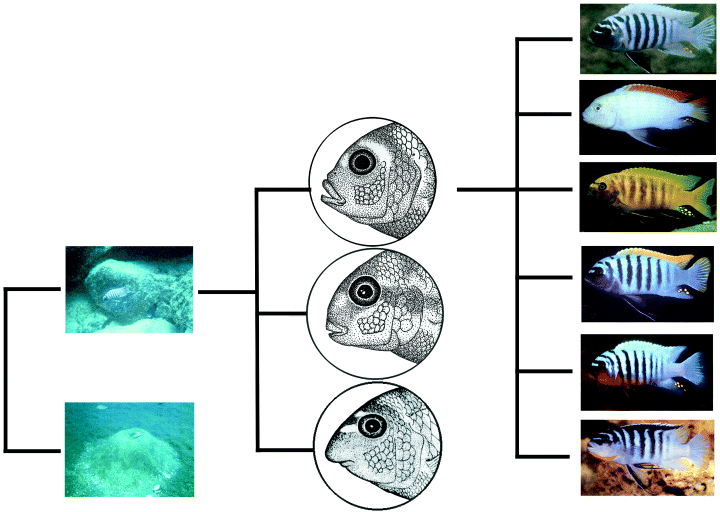
Itt a blogban már közel másfél évtizede is írtam egy hosszabb posztot arról, hogy mi különböztet meg minket főemlős rokonaiktól és amint abban is látszik, természetesen a kulcskérdés szinte mindig az agyra vonatkozik. Hiszen vannak egyéb érdekes vonásaink (pl. relatív szőrtelenségünk), de mégis az agyunk, pontosabban az agyunk által “kódolt” szellemi képességeink azok, amelyek minden jel szerint különlegessé tesznek bennünket – nem természetfelettivé, de mégis egy olyan fajjá, ami a saját eredetén és képességein gondolkodni tud.
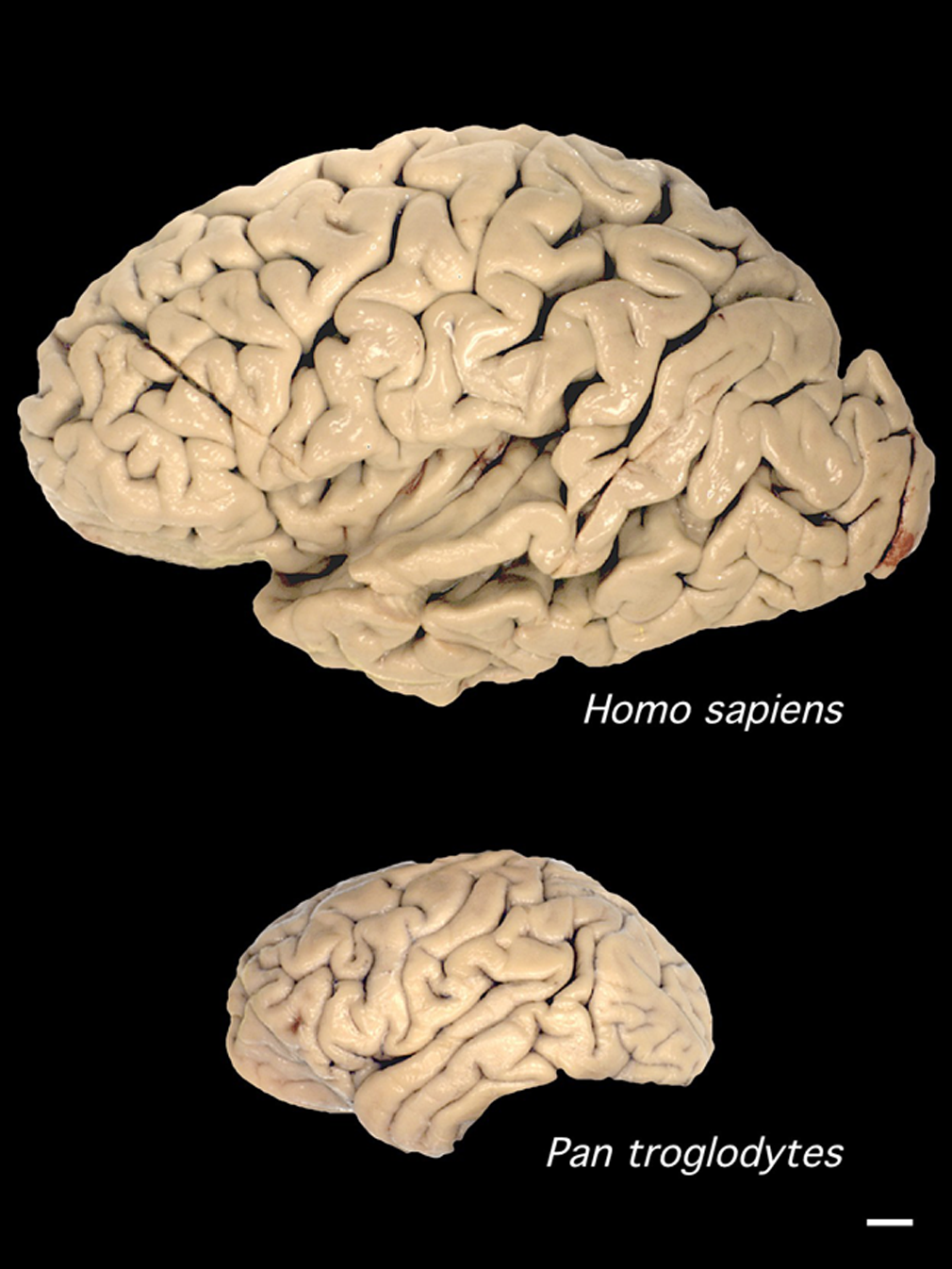
Így aztán minden kor a maga technológiai repertoárjának megfelelően próbált nekiállni az agy vizsgálatának. Amíg csak morfológiát lehetett nézni, addig azt nézték előszeretettel és hasonlították más emlősök, főként emberszabásúak agyának morfológiájához. Amikor később mikroszkópos szintű vizsgálatokra kerülhetett sor, sokan a különböző neuronok morfológiájának, számának és kapcsolatainak feltárásában próbálták keresni a különbségeket. És amikor már genomszekvenciák is elérhetővé váltak, akkor vagy az egyes csimpánz/gorilla/ember szekvencia-szakaszok összevetését vették előszeretettel elő, vagy mindent megtettek, hogy az esetleges különbözőségek biológiai értelmét megfejtsék in vivo kísérletekkel.
Természetesen, mivel emberen és főemlősökön (nagyon helyesen) nem igazán lehet kísérletezni, korábbi technológiák esetében az in vivo kísérlet elsősorban azt jelentette hogy különböző gén-szakaszok emberi és csimpánz megfelelőit fejeztették ki egérben.
Az elmúlt években az in vitro , vagyis “Petri-csészében” nevelt, jellegzetes struktúrát és differenciációs mintázatot mutató 3D-s sejttenyészetek, az organoidok és még konkrétabban az agy organoidok (populárisabb nevükön “miniagyak”) formájában egy teljesen új technológia vált elérhetővé. Ezeknek a miniagyak használata egyébként is érdekes és fontos etikai kérdéseket vet fel, és mivel alapvetően érdekes dolgokat árulhatnak el az idegrendszer korai fejlődéséről, nyilvánvalóan csak idő kérdése volt, hogy valaki felhasználja őket a köztünk és más emberszabásúak közti különbségek boncolgatására.
Ez a “valaki” aztán annak a Madeline Lancasternek a csoportja lett, aki Jürgen Knoblich bécsi laborjában poszt-dokként eredetileg is az úttörője volt az agy organoidok készítésének és tanulmányozásának.
Az új Cell-ben közölt tanulmányban csimpánz és gorilla “miniagyakat” vetettek össze a (most már rutinból készített) emberi agy organoidokkal. Külsőre ezek eléggé hasonló struktúrákat hoztak létre, azzal a különbséggel, hogy az emberi organoidok jellemzőek nagyobbak voltak, mint a másik két emberszabásúé, bennük egy viszonylag nagyobb üreggel.
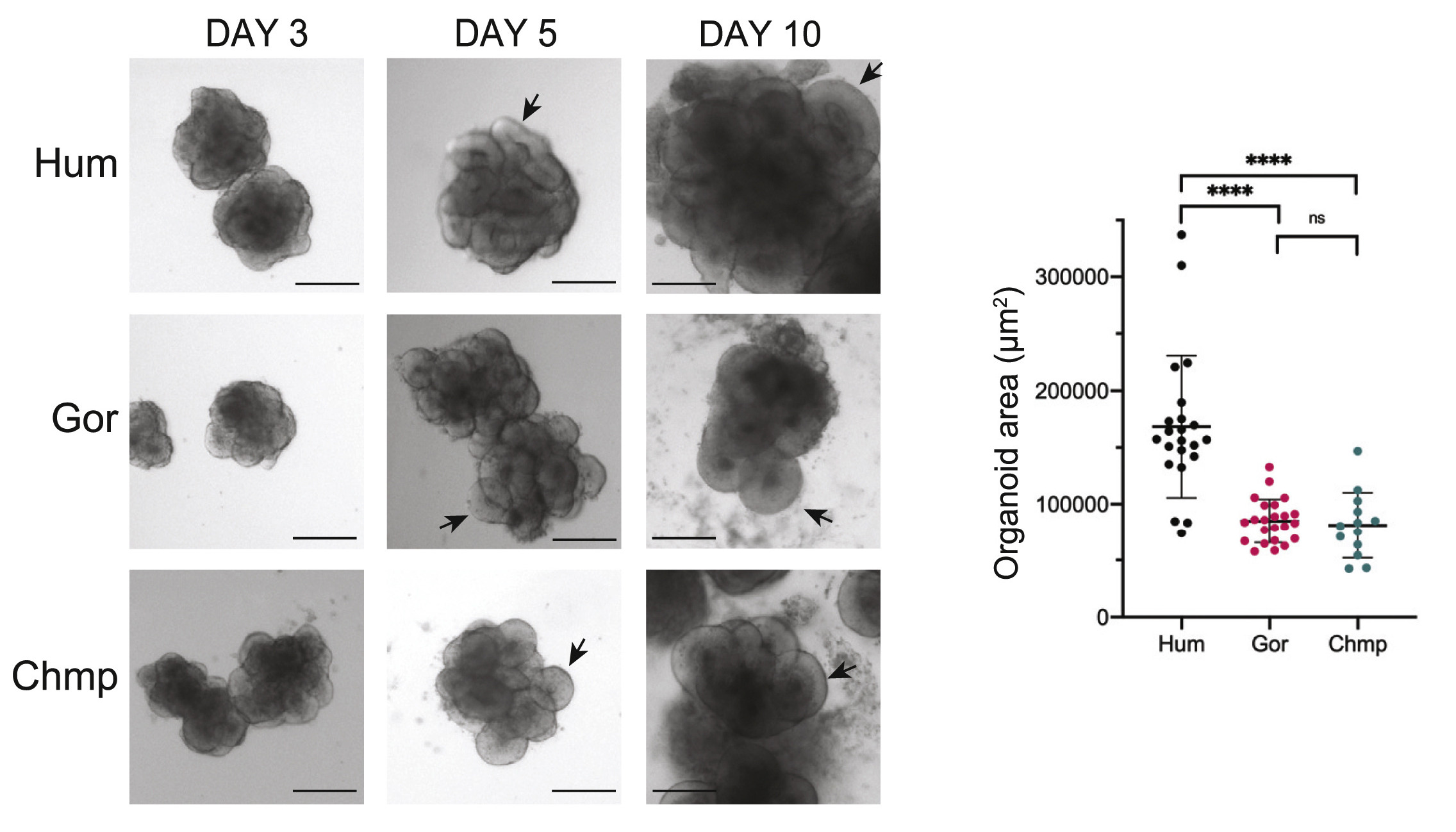
Mindez arra utalt, hogy az emberi organoidokban egységnyi idő alatt több sejt keletkezik. Persze ezt igazolni is kellett, és végül több módszer segítségével ezt meg is tették. Ezeknek a megfigyeléseknek az érdekese hozadéka az a megfigyelés, hogy az emberi organoidokban az idegrendszeri progenitor sejtek gyorsabban osztódnak és később kezdenek el differenciálódni.
Az embrionális fejlődés során is az idegszövet kezdeti, “éretlen” állapota egy olyan ún. neuroepitélium (NE), ahol kicsit elnyúlt sejteket találunk, amelyek a kialakuló szövet mindkét oldalával (apikális, illetve bazális, szakzsargonban) kapcsolatban van. A sejtmagok egy jellegzetes jojó-mozgást végeznek ebben a stádiumban, oda-vissza mozognak a két felszín közt a sejtciklusuk alatt (ún. interkinetikus nukleáris migráció) és az apikális felszín közelében osztódnak.
Az organoidokban is ez ismétlődik meg (az apikális felszín az organoid belsejében nyíló üreg oldalán lesz) és az emberi miniagyakban gyorsabb ez a mozgás és későbbre tolódik az a pillanat, amikor a NE sejtek elkezdenek átalakulni (tNE), ami az első jele annak, hogy megkezdődik az idegszövet differenciációja.

2021-ben nincs már magára valamit is adó cikk rettenetesen sok szekvenálási adat nélkül, így itt is rengeteg stádiumban meghatározták a fejlődő emberi és gorilla miniagyak génexpresszióját, olyan géneket keresve, amelyek egy kicsit más viselkedést mutatnak a két rendszerben.
A kísérlet logikája az volt, hogy feltételezhető, a kicsit eltérő sejtciklusok mögött valami szabályozó génnek a megváltozott expressziója állhat – ami, tegyük hozzá, azért egy nem minden kockázat nélküli hipotézis, főleg, hogy az ilyen típusú “adathorgász” megközelítések rengeteg potenciális lehetőséget szoktak feldobni, amelyek közül nagyon sok egyszerűen a szekvenálási adatokban jelen levő biológiai “zaj” eredménye és nincs igazi biológiai relevanciája. Végül azonban találtak/kiválasztottak egy ZEB2 nevű gént, amit tovább is vizsgáltak.
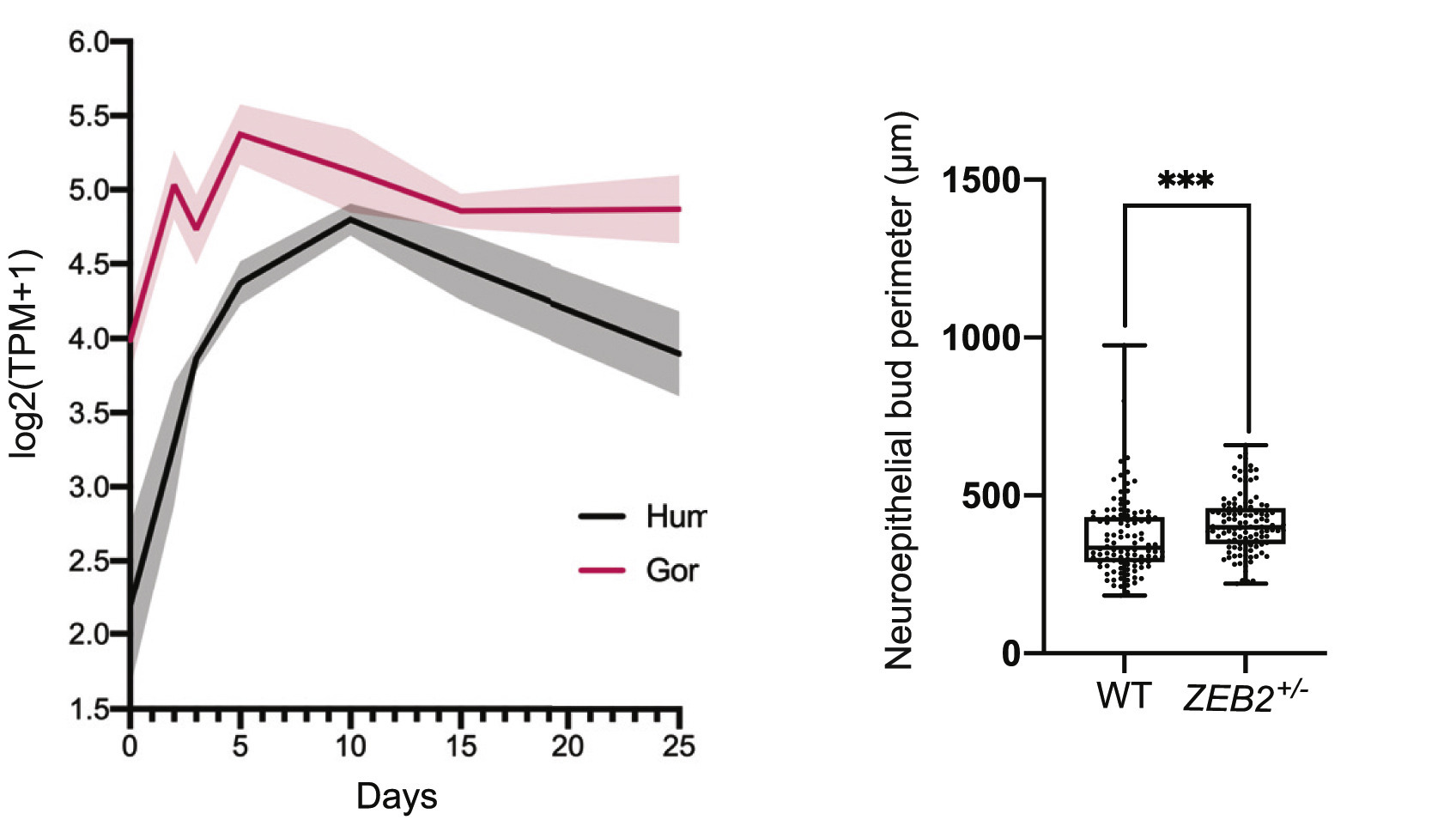
Azért a ZEB2-re esett a választásuk, mert egyrészt ez pont olyan expressziós profillal rendelkezik, hogy egy picit hamarabb van az expressziós csúcsa a gorilla organoidokban, mint az emberben, másrészt maga a gén is számos olyan jelet mutat, hogy az ember felé vezető leszármazási vonalban felgyorsult az evolúciója és pár nagyon jellegezetes aminosav-változást hordoz.
Amikor aztán olyan emberi miniagyakat hoztak létre, amelyekben lecsökkentették a ZEB2 funkcióját, azok kicsit, de azért mérhetően nagyobbak lettek. Hogy ez olyan szignifikáns változás, ami valóban egyértelműen bizonyítja a ZEB2 evolúciós szerepét valószínűleg vitatható lesz: míg a három faj organoidjai közt egyértelmű méretkülönbségek voltak, a ZEB2-hiányos organoid esetében a Lancaster-csoport már csak az organoidok oldalán megjelenő kis kitüremkedések (bud perimether) tudtak valamit kimérni.
A ZEB2 fehérje egyébként BMP-jelátviteli útvonal modulátoraként léphet itt fel és azt szabályozhatja, hogy mennyire hamar kezdődik el az a morfológiai átalakulás az apikális oldalán a NE sejteknek, ami aztán a differenciáció (vagyis a sejtciklusból való kilépés) felét tett egyik első fontos lépés lehet.
Összességében a cikk sok szempontból fontos mérföldkő lehet az ilyen típusú kutatásokban, de ahogy a korábbi hasonló tanulmányok esetében, még azért el kell egy kis időnek telnie, hogy meg tudjuk ítélni, mennyivel kerültünk valóban közelebb az emberi agy különlegességének megértéséhez.
Benito-Kwiecinski S., Giandomenico SL, Sutcliffe M, Riis ES, Freire-Pritchett P, et al. (2021) An early cell shape transition drives evolutionary expansion of the human forebrain. Cell https://doi.org/10.1016/j.cell.2021.02.050.
 Az
Az 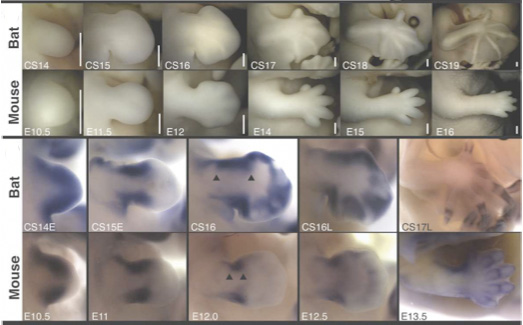
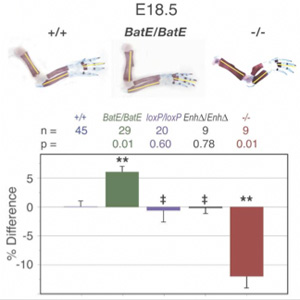

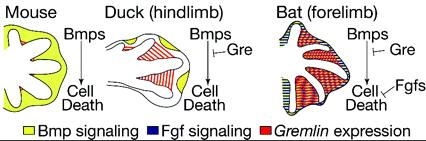


 A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd
A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd