
Utóbbi időben inkább csak szórakozás gyanánt látogatok a magyar kreacion Értelmes Tervezettség Mozgalom (ÉRTEM) honlapjára: ritkán lehet ennyire szépen dokumentálva látni az emberi ignoranciát. A legtöbb fellelhető iromány bevett receptje valahogy úgy kezdődhet: végy egy témát, amiről hibás részinformációid vannak, majd utóbbiakat felhasználva írj valamit, amiben a téma felkent apostolának állítod be magadat.
Első osztályú szintézise ennek a mentalitásnak (egyik személyes kedvencem) Jeszenszky Ferenc (JF) fizikus levele, amiben talán még az ÉRTEM mércéje szerint is durva tévedések vannak (s mivel Jeszenszky a mozgalom egyik tiszteletben álló figurája, próbáljuk meg – vagy inkább ne … – elképzelni, mi van azok fejében, akik számára ő az információforrás…).
Mivel nem vagyok fizikus, maradjunk a biológiai témáknál, bár annyit azért megemlítek, hogy a termodinamika második főtételére vonatkozó érvelése – már, ha jól értelmezem, mert egy kicsit zavaros -, az én szememben még mindig nem áll sziklaszilárdnak mondható lábakon: bármelyik klasszikus kísérletet nézzük, a kiinduló egyszerű anyagokból, bizony komplexebbek jöttek létre (aminosavak, nukleotid bázisok stb).
Sajnos nem tudom a posztban fokozatosan növelni a "feszültséget", ugyanis a csúcs az rögtön a levél elején található. Olyan elképesztő magassága ez az emberi alulinformáltságnak, hogy szinte-szinte már szédítő:
"> 1. Miért tartják olyan
lehetetlennek, hogy a Föld több milliárd éves életkora alatt a
véletlenszerű folyamatok eredményezhettek valami "értelmeset"? Hiszen
elég idő volt rá, és a Földnek is lehet "szerencséje"!” <
Elöljáróban
megjegyezném, hogy (figyelmen kívül hagyva a korokra vonatkozó vitákat)
a szokásos kronológia szerint nem lehet szó több milliárd évről, hiszen
a különböző leletek nemigen engednek meg 100 millió évnél többet az
élet kialakulására."
Jajanyám. Miért pont 100 millió? Miért nem rögtön hatezer, az legalább konzisztens lenne, a klasszikus kreacionista érvekkel? A Föld életkorát 4.5 milliárd évre tesszük és a legkonzervatívabb becslések szerint is 3.2 milliárd éve már léteztek élő szervezetek. Durván 100 millió éve már az első erszényes és méhlepényes emlősök is megjelentek, 200 millió éve (a Jurában) dinoszauruszok folytak a csapból is, 400 millió éve kezdtek a gerincesek kimászni a tengerekből, satöbbi. (Itt lehet gyakorolni.) Ezekután vajon JF milyen alapon várja el, hogy ebben a vitában komolyan vegyék…?
"A morfológia egyetlen olyan esetre sem tud hivatkozni, hogy egy
élőlénynek olyan szerve lenne, amely valamilyen teljesen új életforma
kezdeménye lenne. A drosophila melanogasterrel végzett
évszázados kísérletek azt mutatják, hogy a generációk ezrein keresztül
az erőltetett mutációk sem hoztak létre mást, mint az alapfaj
torzszülötteit."
Rágjuk ezt itt egy picit:"A morfológia egyetlen olyan esetre sem tud hivatkozni, hogy élőlénynek olyan szerve lenne, amely valamilyen teljesen új életforma
kezdeménye lenne". Ha a magyar nyelv szabályai szerint értelmezzük ezt a kis mondatot, kiemelve, különállóan, akkor gondolhatnánk arra, hogy valami olyasmire gondol az írója, mint a hidrák aszexuális szaporodása során megjelenő bimbók, amelyek egy későbbi, a szülővel genetikailag megegyező hidra kezdeményei. Ekkor ugyan nyilvánvalóan nem lenne igaz, de legalább lenne valami értelme.
Amit JF mondani szeretne (és amit a kiemelt mondat egyáltalán nem jelent) az feltehetőleg valami olyasmi, hogy szerinte nem ismert, miként jelennek meg komplexebb állatcsoportok bélyegei az evolúció során. Például halakban a lábak – de nézd, hopp, ilyen is van. Ejnye, akkor ez sem igaz. Arról az inszinuációról nem is szólva, hogy mivel száz év alatt nem sikerült muslicából rozmárt mutáltatni, nem működik az evolúció.
"Arról elgondolkodhatunk, hogy ezek valóban biológiai hipotézisek-e.
Hajlok arra a véleményre, hogy nem. Ugyanis semmiféle valódi tudományos
tény nem támasztja alá egyiket sem. Azonban itt egy tudománytörténeti
problémába ütközünk. Az evolúcióelmélet nem úgy jött
létre, ahogyan általában a tudományos elméletek (mint mondjuk az
elektrodinamika Maxwell-féle elmélete), hogy a megfigyelt tények
kikényszerítenek egy elméleti magyarázatot; hanem úgy, hogy először
volt egy ideológiai alap (nevezetesen, hogy a természet önmagában
megmagyarázható, transzcendens tényezők pedig nincsenek), és ehhez
próbálták igazítani az elméletet."
Egy: iszonyúan nagy ismeretanyag támasztja alá az evolúciót – fejlődésbiológiai, paleontológiai, geográfiai, genetikai kísérletek ezrei. Most ezek JF szerint valójában nem "tudományos tények", vagy egyszerűen nem is tud róluk. Kettő: de bizony, éppen úgy jött létre, hogy a megfigyelések kikényszerítettek egy új elméleti magyarázatot. Darwin idején már több eresztékében recsegett-ropogott a kreacionizmus kártyavára: a halmozódó fosszilis leletanyag nyilvánvalóvá tette, hogy a Földön az élet változik. Nem is említve, hogy a vallás és az evolúció összeférhetetlensége, csak a kreacionisták fantáziájában létezik. JF állításának egyszerűen semmi, de semmi alapja nincs.

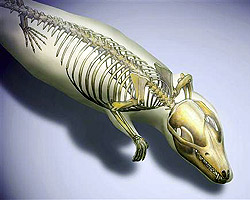 A kínai lelőhelyek továbbra is ontják magukból az érdekesnél érdekesebb fosszíliákat. Most éppen ismét egy ősi emlős került elő, amely egyszerre támasztja alá a középfül evolúciós kialakulásáról szóló elméleteinket és kínál betekintést az evolúció boszorkánykonyhájának egy Hox-gének tüzelte bugyrába.
A kínai lelőhelyek továbbra is ontják magukból az érdekesnél érdekesebb fosszíliákat. Most éppen ismét egy ősi emlős került elő, amely egyszerre támasztja alá a középfül evolúciós kialakulásáról szóló elméleteinket és kínál betekintést az evolúció boszorkánykonyhájának egy Hox-gének tüzelte bugyrába.
 A napfény egyszerre jelent áldást és gondforrást: utóbbit a benne levő
A napfény egyszerre jelent áldást és gondforrást: utóbbit a benne levő  A malária elleni védekezésről beszélve, általában hajlamosak vagyunk egy kalap alá venni magát a malária parazitát és köztes gazdáját, a szúnyogot: a cél mindkettő pusztítása. Persze ez nem meglepő, hiszen a gyakorlatban is elég jól helytáll a "kevesebb szúnyog = kevesebb megbetegedés" egyenlet. De ennek fényében gyakran elfeledkezünk egy apróságról: a Plasmodium a szúnyogoknak is a parazitája, nem csak nekünk, embereknek.
A malária elleni védekezésről beszélve, általában hajlamosak vagyunk egy kalap alá venni magát a malária parazitát és köztes gazdáját, a szúnyogot: a cél mindkettő pusztítása. Persze ez nem meglepő, hiszen a gyakorlatban is elég jól helytáll a "kevesebb szúnyog = kevesebb megbetegedés" egyenlet. De ennek fényében gyakran elfeledkezünk egy apróságról: a Plasmodium a szúnyogoknak is a parazitája, nem csak nekünk, embereknek.
 A
A  Az egerek világa, a szó szoros értelmében, lényegesen színtelenebb, mint a miénk, hiszen az
Az egerek világa, a szó szoros értelmében, lényegesen színtelenebb, mint a miénk, hiszen az  A
A 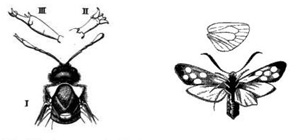 Ha szemügyre veszünk egy ezerlábút, talán az első dolog ami a szemünkbe ötlik, hogy teste ismétlődő egységekből (ún. szegmensekből) áll, amelyek mint megannyi egyforma „építőmodul” sorakoznak egymás mögött. Figyelmesebben szétnézve valami hasonlóra lelhetünk rákokban, rovarokban és más ízeltlábúakban is, sőt, mi magunk, gerincesek sem vagyunk kivételek az efajta modularitás alól, bár esetünkben az ismétlődő egységeket leginkább csak az embrionális fejlődés során lehet könnyen elkülöníteni. S ugyan fontos hangsúlyozni, hogy utóbbi állatcsoportokban, az ezerlábúak szegmenseitől eltérően, az „építőmodulok” nem tökéletesen egyformák, mégis egyértelműnek tűnik, hogy amit látunk az egyfajta „variációk egy témára”, vagyis minden egység leszármaztatható egy szegmens-prototípusból. Ebből persze rögtön adódik a kérdés: milyen mechanizmusok felelősek az egyes szegmens-variánsok kialakulásáért?
Ha szemügyre veszünk egy ezerlábút, talán az első dolog ami a szemünkbe ötlik, hogy teste ismétlődő egységekből (ún. szegmensekből) áll, amelyek mint megannyi egyforma „építőmodul” sorakoznak egymás mögött. Figyelmesebben szétnézve valami hasonlóra lelhetünk rákokban, rovarokban és más ízeltlábúakban is, sőt, mi magunk, gerincesek sem vagyunk kivételek az efajta modularitás alól, bár esetünkben az ismétlődő egységeket leginkább csak az embrionális fejlődés során lehet könnyen elkülöníteni. S ugyan fontos hangsúlyozni, hogy utóbbi állatcsoportokban, az ezerlábúak szegmenseitől eltérően, az „építőmodulok” nem tökéletesen egyformák, mégis egyértelműnek tűnik, hogy amit látunk az egyfajta „variációk egy témára”, vagyis minden egység leszármaztatható egy szegmens-prototípusból. Ebből persze rögtön adódik a kérdés: milyen mechanizmusok felelősek az egyes szegmens-variánsok kialakulásáért?  A
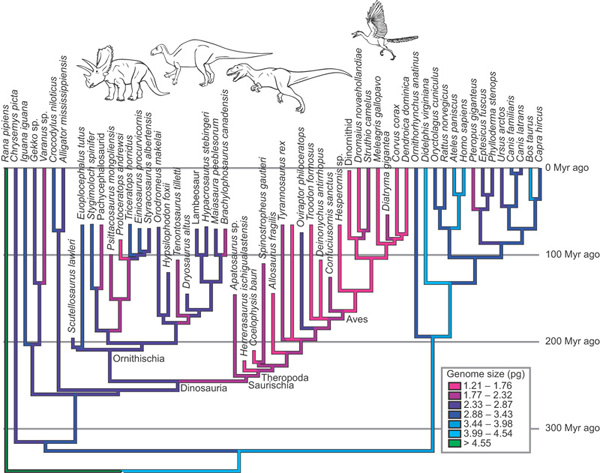
A  A testmérethez viszonyított könnyű súly nem éppen hátrányos a repülésnél, ezért alakult ki a madarak üreges csontozata is. Ha azonban elfogadunk egy, a Nature-ben publikált új elméletet, a fizikai fölöslegtől való megszabadulás csak az egyik feltétele a levegő meghódításának. Egy másik, a felesleges DNS, azaz a nem kódoló genetikai anyag kiszelektálása lehet. (A "nem kódoló" itt szélesebb értelemben veendő, és nemcsak a fehérjék aminosav-sorrendjének leírására, hanem az egyes gének szabályozó régióira is vonatkozik.)
A testmérethez viszonyított könnyű súly nem éppen hátrányos a repülésnél, ezért alakult ki a madarak üreges csontozata is. Ha azonban elfogadunk egy, a Nature-ben publikált új elméletet, a fizikai fölöslegtől való megszabadulás csak az egyik feltétele a levegő meghódításának. Egy másik, a felesleges DNS, azaz a nem kódoló genetikai anyag kiszelektálása lehet. (A "nem kódoló" itt szélesebb értelemben veendő, és nemcsak a fehérjék aminosav-sorrendjének leírására, hanem az egyes gének szabályozó régióira is vonatkozik.)
{kind=link}
{kind=link}