Ezúttal mellőzve az ilyenkor kötelező mentegetőzési kört a posztok megritkulása/nemléte okán, csapjunk is bele, miket olvastunk az elmúlt évben:
Könyvhét – 2024
Válasz

Ezúttal mellőzve az ilyenkor kötelező mentegetőzési kört a posztok megritkulása/nemléte okán, csapjunk is bele, miket olvastunk az elmúlt évben:

Kevés dolog vált annyira evidenciává a biológia tankönyvek evolúcióról szóló fejezetében, mint a pikkelyek és tollak (és persze a szőr) homológiája. A madarak hüllő-rokonságának egyre pontosabb feltárása, a tollas dínókra való rácsodálkozás az elmúlt évtizedben mind-mind segített ezt a homológiát ismertté tenni. Ennek megfelelően számos tankönyv és renegeteg összefoglaló tanulmány illusztrálta, hogy miként is alakul(hatot)t ki az egyszeri pikkelyből toll az evolúció folyamán.
És épp ezért volt bosszantó, hogy egészen a közelmúltig a legfontosabb bizonyíték mégiscsak hiányzott a témában: az, hogy a fejlődés során hasonló képletekből alakulnak ki a későbbi pikkelyek, illetve tollak.
Tízéves szülinapi bulink másik előadója, Lippai Mónika (ELTE TTK, Anatómiai, Sejt- és Fejlődésbiológiai Tanszék), a β-importin fehérje sokoldalú funkcióiról, illetve azok felfedezéséről mesélt. (Köszönet Tóth Andrásnak a videóért.)
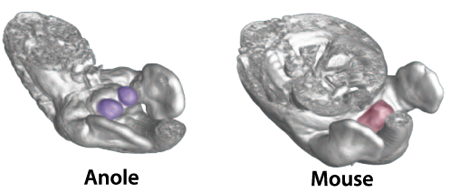 A szárazföld meghódítása nemcsak az úszás helyett mászáshoz alkalmazkodott végtagokat igényelt, de a korábbi szaporodási stratégiák radikális újragondolását is szükségessé tette a gerincesekben.
A szárazföld meghódítása nemcsak az úszás helyett mászáshoz alkalmazkodott végtagokat igényelt, de a korábbi szaporodási stratégiák radikális újragondolását is szükségessé tette a gerincesekben.
Hiszen míg a vizes közegben az ivarsejtek szabadon engedésével le lehetett tudni az evolúciós sikerhez szükséges hajcihőt, addig a szárazföldön valami közvetlenebb módon kellett célba juttatni őket. És ez a közvetlenebb mód lett a külső ivarszervek kialakulása, amely hímek esetében a péniszt jelenti.
 Bő éve már írtam egy hosszabb posztot arról, hogy miként magyarázható az ujjaink kialakulása a fejlődés során Turing-mechanizmus segítségével. Akkor valahol ott hagytuk abba, hogy minden jel szerint ha létezik egy ilyen mechanizmus, azt Hox fehérjék (elsősorban Hoxd13), illetve a Fibroblast Growth Factor (Fgf) jelátvitel modulálja.
Bő éve már írtam egy hosszabb posztot arról, hogy miként magyarázható az ujjaink kialakulása a fejlődés során Turing-mechanizmus segítségével. Akkor valahol ott hagytuk abba, hogy minden jel szerint ha létezik egy ilyen mechanizmus, azt Hox fehérjék (elsősorban Hoxd13), illetve a Fibroblast Growth Factor (Fgf) jelátvitel modulálja.
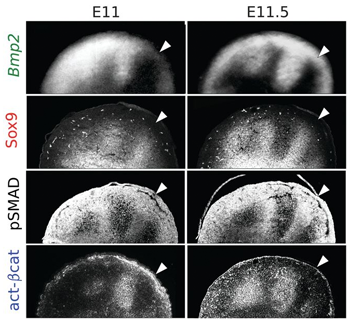
Most a korábbi tanulmány szerzői közül James Sharpe csoportja tovább igyekezett göngyölíteni a rejtélyt és azonosították a korábban csak feltételezett Turing-mechanizmus molekuláris komponenseit. Ezek pedig nem mások, mint két másik fontos jelátviteli útvonal a Bmp és Wnt elemei, valamint a csontképződés egyik legkoraibb markerének tartott Sox9.
Az már korábban is ismert volt, hogy a Bmp jelátvitel elősegíti a Sox9 kifejeződését, a Wnt jelátvitel pedig gátolja azt, de Sharpejék először figyeltek fel rá, hogy a Sox9 valamint a Wnt jelátvitel (aktivált B-catenin által mért) aktivitása és a Bmp jelátvitel egyik ligandumát kódoló Bmp2 gén expressziója szintén komplementer. (Érdekes módon a Bmp2 kifejeződése, és a Bmp jelátvitel (pSMAD-al mérhető) aktivitása sem esik egybe, ami annak az oka, hogy ahol a ligandum aktiválni tudja a receptort ott egy ún. feed-back gátlás valósul meg a bekapcsolt gének, így a Sox9 által is.)
 A béka hátsó lábában, a gekkók melső végtagjában és a mi kezünkben a számtalan homológ csontocska mellett is feltünő egy, fontos, közös vonás: mindannyiunknak öt ujja van. Az öt ujjúság a szárazföldi négylábú gerincesek nagy közös vonása, olyannyira, hogy egy tudományos kifejezés is születtet a sztereotip négylábú végtag leírására: pentadactyl végtag.
A béka hátsó lábában, a gekkók melső végtagjában és a mi kezünkben a számtalan homológ csontocska mellett is feltünő egy, fontos, közös vonás: mindannyiunknak öt ujja van. Az öt ujjúság a szárazföldi négylábú gerincesek nagy közös vonása, olyannyira, hogy egy tudományos kifejezés is születtet a sztereotip négylábú végtag leírására: pentadactyl végtag.
Az ettől való eltérés mindig különös és az evolúciósan csak egy irányba történik: a békák melső lábán, a madarak szárnyában, vagy a lovak patájában egyaránt kevesebb ujj figyelhető meg (négy, három, illetve egy). Az ötnél több ujjúság (az ún. polydactylia) kóros esetekben észlelhető, például a végtagok fejlődése apropóján korábban már emlegetett Sonic-hedgehog (Shh) gén mutációja okozhat ilyen elváltozást.
De vajon miért pont öt a normális, illetve lehette-e valamikor hat, vagy hét? Esetleg tizenkettő?
 Sajnos az utóbbi napok oroszlán-központúsága elnyomta a Budapesti Állatkert igazán fontos új lakóinak érkezését: márpedig a Varázshegy sötétlabirintusában otthont lelő két nagy ugróegér (Jaculus orientalis) lényegesen több figyelmet érdemelne.
Sajnos az utóbbi napok oroszlán-központúsága elnyomta a Budapesti Állatkert igazán fontos új lakóinak érkezését: márpedig a Varázshegy sötétlabirintusában otthont lelő két nagy ugróegér (Jaculus orientalis) lényegesen több figyelmet érdemelne.
Egy átlagos Pokémon-figura valószínűtlen testalkatával rendelkező állatok amellett hogy zoológiai különlegességek, egy fejlődésbiológus számára igazi sztárok, hiszen – akárcsak egy denevér esetében – embriológiájuk megértése sok mindent elárul a kevésbé különleges anatómiával rendelkező emlősök fejlődéséről is.
Mint azt a szex determinációról pár hónapja született posztban is olvashattuk, egy adott egyedben a nemet meghatározó kromoszómák lehetnek különbözőek (XY), illetve azonosak (XX). Ez utóbbi esetben az emlősöknél fellép egy aprócska probléma: a két X kromoszóma kétszer annyi génterméket eredményez női ivarú egyedekben, mint hímekben. Vagyis csak eredményezne, létezik ugyanis egy igen érdekes jelenség, a dóziskompenzáció, ami az egyik X kromoszómát – génexpressziós szempontból – elnémítja.
Az egyik X kromoszóma kikapcsolása az emlős egyedfejlődés során igencsak korán, pársejtes állapotban minden egyes sejtben külön végbemegy, és ennek a mintázata a soron következő mitózisok során tovább adódik. Mivel a sejtekben véletlenszerűen vagy az apai vagy az anyai X kromoszóma inaktiválódott, ezen sejtek későbbi utódsejtjeiben értelemszerűen vagy az apai vagy az anyai X kromoszómán található génvariánsok fejeződnek ki – esetleges mozaikos mintázatot eredményezve (lásd a teknőctarka macskák esetét).
 Amikor egyszer régen írtam egy hosszú, bevezető posztot az emlős végtagok fejlődéséről, akkor az volt a nem is igen titkolt szándékom, hogy az egész amolyan felvezetésként szolgál majd a mellső végtag evolúcióját tárgyaló posztokhoz. Aztán teltek az évek és igazából csak nem sikerült időt szakítani a témára, pedig mindig lett volna anyag. Múlt hónapban azonban két olyan fontos cikk is kijött, ami miatt most mégis össze kell kapnom magam, nem is egy, hanem két poszt erejéig, mert ezekről hallgatni már-már bűn lenne. 🙂
Amikor egyszer régen írtam egy hosszú, bevezető posztot az emlős végtagok fejlődéséről, akkor az volt a nem is igen titkolt szándékom, hogy az egész amolyan felvezetésként szolgál majd a mellső végtag evolúcióját tárgyaló posztokhoz. Aztán teltek az évek és igazából csak nem sikerült időt szakítani a témára, pedig mindig lett volna anyag. Múlt hónapban azonban két olyan fontos cikk is kijött, ami miatt most mégis össze kell kapnom magam, nem is egy, hanem két poszt erejéig, mert ezekről hallgatni már-már bűn lenne. 🙂
Az első cikk azzal foglalkozik, hogy miképpen befolyásol(hat)ta az elnyújtott Hoxd13 expresszió a végtagok fejlődését. De, hogy ne rontsunk ajtóstól a házba, előbb lássuk, hogyan kerül elő egyáltalán ez a gén és egyáltalán, miben is különbözik egy egér és egy hal végtagja.
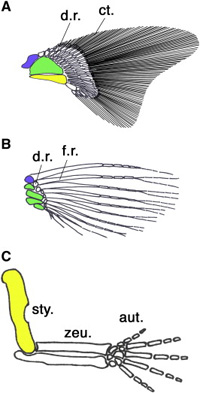
Ha az ősi gerinces végtagot vesszük szemügyre, ami ma is megfigyelhető egyes procoshalakban (lásd cápák – A), akkor azt látjuk, hogy a végtag ízesülését a vállövhöz három nagy csont biztosítja, ezek a propterygium (kék), mesopterygium (zöld) és metapterygium (sárga). Ezekhez apróbb csontok kapcsolódnak (distal radials – d.r.), amelyek az úszósugarakat tartják. Csontoshalak esetében (B) ez a szabásminta úgy egyszerűsödött, hogy a metapterygium elvesződött, és az úszósugarakat tartó apróbb csontok száma is redukálódott. A szárazföldi gerincesekben (C) viszont pont a metapterygium maradt egyedül meg (felkarcsontként), a disztális radiálisokból pedig kialakultak az alkarunk csontjai és a kéztőcsontok. (Némi nemű vita van arról, hogy az ujjak maguk az úszósugarak homológjai-e, vagy sem, de a jelenleg dominánsabbnak tűnő nézet szerint igen.)
A hét egyik tudományos bulvárszenzációja a Mexikóban lelt egyszemű cápa-bébi lett, amit sikerült mindenütt (így az elvileg tudományos portálként definiálható LiveScience-n is) elképesztően blőd körítéssel tálalni.
Pedig a jelenség, ami a szerencsétlen kis lényt minden valószínűség szerint ilyenné tette, kialakítva a holoprosencephalia tünetegyüttest, egy elég univerzális gerinces fejlődési útvonal elromlása lehet (ami összességében nem is annyira hallatlanul ritka, lásd nem is olyan régen Cy, a küklopsz-kismacska esetét).
