 Mint pár hete a Xiphophorus nemzettségbe tartozó halak, és a bennük levő onkogén kapcsán szóba került, a valóság néha kínál olyan eseteket, amikor a természetes szelekció és a szexuális szelekció egymással ellentétes éredekeket képviselnek.
Mint pár hete a Xiphophorus nemzettségbe tartozó halak, és a bennük levő onkogén kapcsán szóba került, a valóság néha kínál olyan eseteket, amikor a természetes szelekció és a szexuális szelekció egymással ellentétes éredekeket képviselnek.
Azonban ez egyáltalán nem törvényszerű, s gyakran a szexuális szelekció történetesen rájátszik a természetes szelekció által favorizált változásokra. Hogy erre jó példát mutassunk, elég egy másik halnemzettségre tekintenünk.
A Viktória-tóban élő bölcsőszájú halakról van szó, pontosabban azok közül is a Pundamilia fajokról. Ezek a halak a part közelében élnek, viszonylag átfedő életterekben (szakszóval élve szimpatrikus fajok), és bár a fajok tipikus egyedei könnyen elkülöníthetőek, a kinézeti-szórás elég nagy, és lesznek olyan egyedek, amelyekről pusztán kinézet alapján nehéz megmondani, hogy hova is tartoznak.
A baloldali képen a P. pundamilia és a P. nyererei „nászruhába” öltözött tipikus hím egyedei láthatóak, felül illetve alul, valamint a kevésbé tipikus hímek (a félreértések elkerülése végett: ezek nem hibridek, hanem nagyon is az egyik, vagy a másik fajhoz tartoznak, molekuláris szinten ez azonosítható). A P. nyererei egyébként átlagosan a mélyebb vizeket kedveli, és ez adta az ötletet a kutatóknak, hogy egy kézenfekvő hipotézist teszteljenek: mivel a Viktória-tóban, a mélység növekedésével arányosan egyre csökken a lejutó fény kék komponensének a mennyisége – ez a vízben úszkáló részecskéknek köszönhető – (vagyis relatíven a piros komponens mennyisége nő), elképzelhető, hogy a tipikusan itt élő halak esetében a fényérzékelésért felelős opszin fehérjéket kódoló gének úgy változtak, hogy a halak látásának érzékenysége eltolódott ennek a hosszabb hullámhosszú (piros) színtartománynak az irányába. Ez pedig hosszabb távon a hímek nászöltözetét is megváltoztatta, hiszen ha a nőstények jobban látják a pirosat, akkor jobban megéri ilyen színekben pompázni előttük.
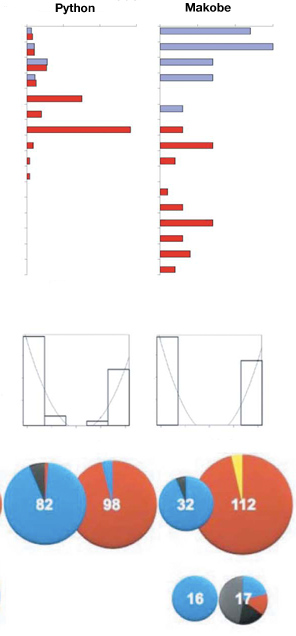 Hogy az elméletet teszteljék, közel ezer hímet vizsgáltak meg a Viktória-tó néhány szigete közelében. Az eredmény elég meggyőző. Először is, különösen a fokozatosan mélyülő helyeken, jól megfigyelhető, mint adja át a helyét a fémes-kék forma, a piros színűnek, a parttól egyre távolodva. Ha elég fokozatos a mélyülés (pl. Makobe estében – a függőleges tengely a mélységet jelöli), szinte nincs is átmeneti forma, csak a két „szélső érték” fedezhető fel, jól elkülönülve . (Ezt hivatottak ábrázolni a fekete-fehér oszlopdiagrammok a második sorban.)
Hogy az elméletet teszteljék, közel ezer hímet vizsgáltak meg a Viktória-tó néhány szigete közelében. Az eredmény elég meggyőző. Először is, különösen a fokozatosan mélyülő helyeken, jól megfigyelhető, mint adja át a helyét a fémes-kék forma, a piros színűnek, a parttól egyre távolodva. Ha elég fokozatos a mélyülés (pl. Makobe estében – a függőleges tengely a mélységet jelöli), szinte nincs is átmeneti forma, csak a két „szélső érték” fedezhető fel, jól elkülönülve . (Ezt hivatottak ábrázolni a fekete-fehér oszlopdiagrammok a második sorban.)
Ha pedig ezeknek a halaknak az opszin génjeit is górcső alá vesszük akkor azt láthatjuk, hogy a hosszú hullámhosszú fény érzékeléséért felelős ún. LWS opszin gén esetében a kék színű populációikban egy kék-színre érzékenyebb allél az elterjedtebb, míg a vörös nászruhás halak esetében a piros színre érzékenyebb allélok (lásd harmadik sor, kördiagrammok).
A Makobe sziget melletti populációk esetében, egy másik opszin gén, a rövid hullámhossz érzékelésében szerepet játszó SWS2 is hasonlóan változott (ezt az utolsó sorban látható, kisebb kördiagrammok mutatják – egyébként a számok, azt jelölik, hogy adott gént hány egyedben szekvenálták meg): a part menti populációban kizárólag a kék színre érzéken allélja van jelen, míg a mélyebb területeken élő, egyedekben már olyan allélikus variánsok is megjelentek, amelyek a vörös színre érzékenyebbek (a fekete és szürke, az „más” allélvariánsokat jelöl – se nem tipikusan „kék-érzékeny”, se nem tipikusan „piros-érzékeny”).
Jól látható tehát, hogy a fokozatosan mélyebb vizek kolonizálásával párhuzamosan zajló speciáció egy várható (és hasznos) élettani változással járt: a szem más hullámhosszra való „hangolásával”. Ez azonban, egy olyan fajban, ahol a hímek nászöltözete kulcsfontosságú reprodukciós sikere szempontjából, szinte „menetrendszerűen” magával hozta a hímek színének megváltozását is.
Ha teljesen őszinték vagyunk, az eredmény azért nem sokkolóan újszerű: szűk két éve ugyanez a csoport valami nagyon hasonlót írt le két másik faj esetében – bár ott a mélység helyett a víz zavarossága volt az opszin gének és a nászmintázat evolúcióját hajtó erő.
Seehausen O, Terai Y, Magalhaes IS, Carleton KL, Mrosso HD, et al. (2008) Speciation through sensory drive in cichlid fish. Nature 455: 620-626.


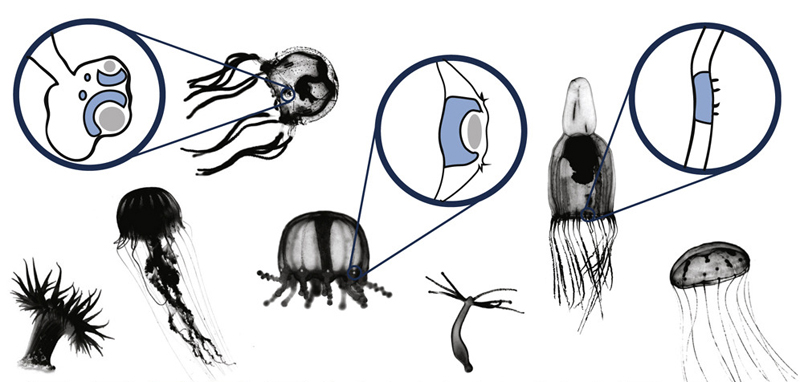
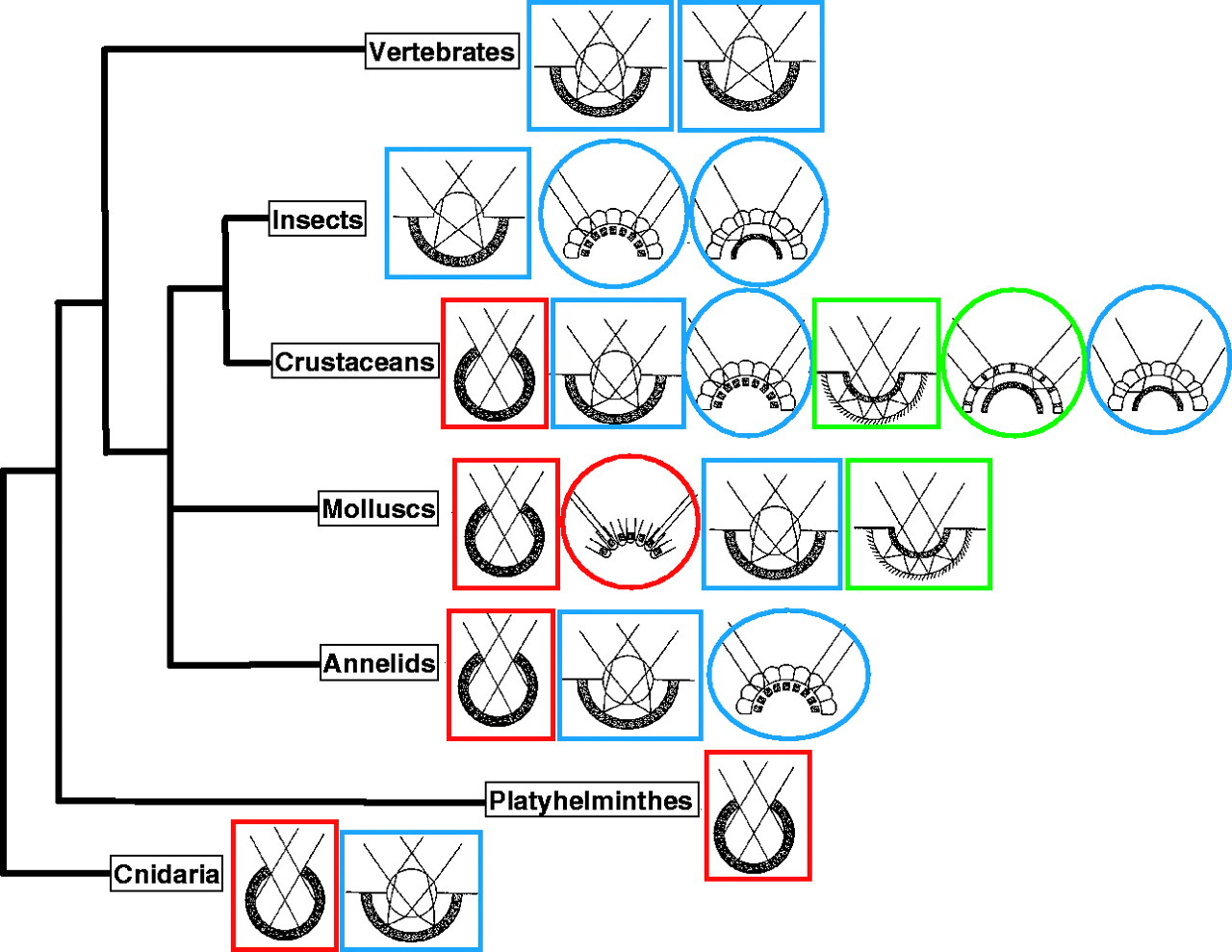
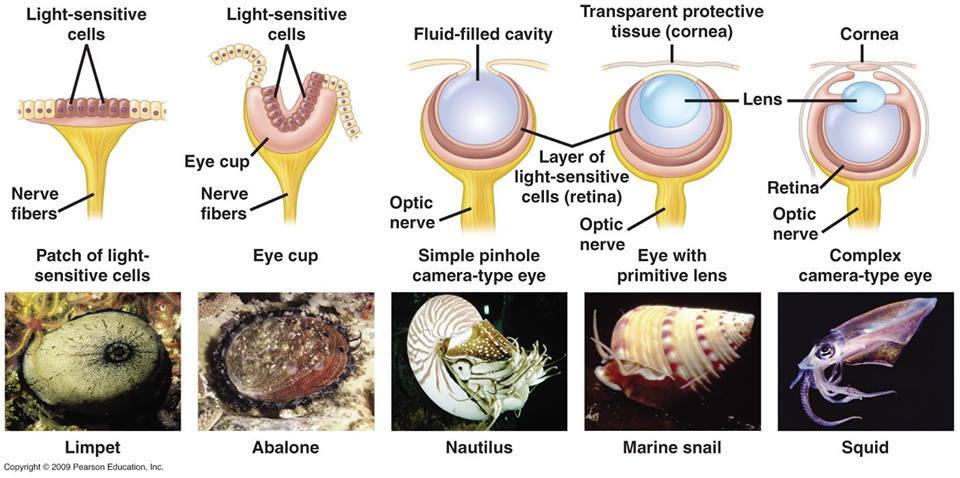
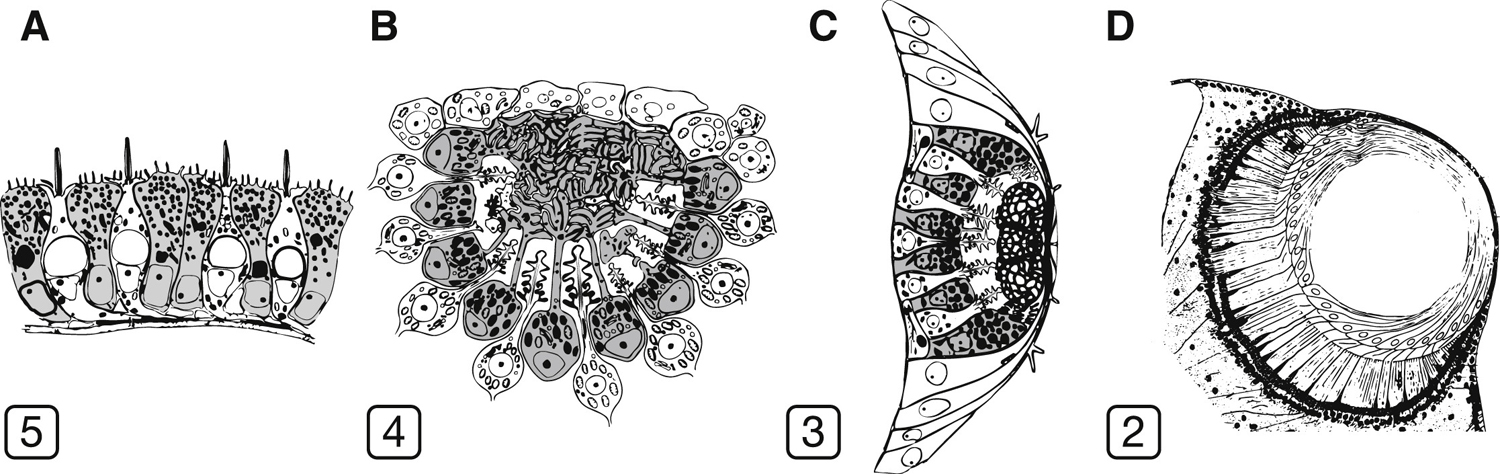

 Méltatlanul régen nem foglalkoztunk a blog egyik kedvenc témájával, a különleges szemekkel, pedig ha egy kicsit is vízközelbe megyünk csak úgy hemzsegnek az egzotikusnál egzotikusabb látószervek:
Méltatlanul régen nem foglalkoztunk a blog egyik kedvenc témájával, a különleges szemekkel, pedig ha egy kicsit is vízközelbe megyünk csak úgy hemzsegnek az egzotikusnál egzotikusabb látószervek: 
 A szem evolúciója hálás téma, a különleges szemeké pedig duplán az. Írtunk már ennek apropóján
A szem evolúciója hálás téma, a különleges szemeké pedig duplán az. Írtunk már ennek apropóján  Mint arról bárki, aki vaksötétben próbált megtalálni egy kulcslyukat tanúskodhat, az éjszakai látás trükkös dolog. A hangsúly egyáltalán nem színeken van, hanem árnylatokon; valami észrevétele és meg nem látása közt alig néhány fotonnyi különbség van.
Mint arról bárki, aki vaksötétben próbált megtalálni egy kulcslyukat tanúskodhat, az éjszakai látás trükkös dolog. A hangsúly egyáltalán nem színeken van, hanem árnylatokon; valami észrevétele és meg nem látása közt alig néhány fotonnyi különbség van.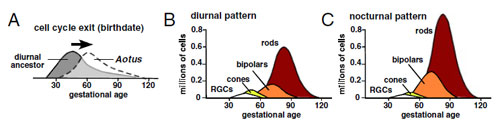 A jelek szerint az Aotus éjimajmok épp ezt a megoldást választották. (Persze ezt kimutatni egy kicsit trükkös, hiszen vizsgált, viszonylag közeli rokon majmok fejlődése is különbözik annyira, hogy nem hasonlítható össze csak a megtermékenyítéstől eltelt idő alapján.) A retina differenciácója az éjszakai fajokban (nocturnal) későbbre tolódott, és ez pont a számukra igen hasznos fényérzékeny sejttípus mennyiségét növelte meg.
A jelek szerint az Aotus éjimajmok épp ezt a megoldást választották. (Persze ezt kimutatni egy kicsit trükkös, hiszen vizsgált, viszonylag közeli rokon majmok fejlődése is különbözik annyira, hogy nem hasonlítható össze csak a megtermékenyítéstől eltelt idő alapján.) A retina differenciácója az éjszakai fajokban (nocturnal) későbbre tolódott, és ez pont a számukra igen hasznos fényérzékeny sejttípus mennyiségét növelte meg.  Vannak olyan pillanatok, amikor egy addig számára ismeretlen fajt látva egy biológus szeme is nagyra tágul, és a hoaxra meg photoshopra gyanakodva kezd keresgélni az eredeti cikk után. Kb. ez volt esetemben is, amikor először megláttam a Macropinna microstoma fényképét.
Vannak olyan pillanatok, amikor egy addig számára ismeretlen fajt látva egy biológus szeme is nagyra tágul, és a hoaxra meg photoshopra gyanakodva kezd keresgélni az eredeti cikk után. Kb. ez volt esetemben is, amikor először megláttam a Macropinna microstoma fényképét. Előre látott és kevésbé látott elfoglaltságok miatt, kicsit poszt-szegényebb időknek néz elébe a blog. De türelem, mert előbb vagy utóbb, visszatérünk a régi rendszereséghez. 🙂
Előre látott és kevésbé látott elfoglaltságok miatt, kicsit poszt-szegényebb időknek néz elébe a blog. De türelem, mert előbb vagy utóbb, visszatérünk a régi rendszereséghez. 🙂