Ezúttal mellőzve az ilyenkor kötelező mentegetőzési kört a posztok megritkulása/nemléte okán, csapjunk is bele, miket olvastunk az elmúlt évben:
Könyvhét – 2024
Válasz

Ezúttal mellőzve az ilyenkor kötelező mentegetőzési kört a posztok megritkulása/nemléte okán, csapjunk is bele, miket olvastunk az elmúlt évben:

Ahogy azt egyszer már pár éve megállapítottam: ha semmi nincs, könyvheti ajánló-poszt akkor is van (na jó, tavaly az is a Könyvfesztiválra csúszott, de az még talán pont belefért). Idén viszont tényleg a 94. Ünnepi Könyvhétre időzítve posztolunk, szóval lássuk mit olvastunk az utóbbi évben, amit ajánlani is tudunk, merünk, vagy épp ellenkezőleg, mindenkit lebeszélnénk róla.
Kalmár Éva
 A Stanford- , a Milgram-, avagy a Tuskagee szifilisz kisérletet ma biztosan nem engedélyezné egyik egyetem sem, mint ahogy amit Dr Murray művelt 1960-ban a fiatal Theodore Kaczynskivel, akiből a hírhedt Unabomber lett, ma már megengedhetetlen: a kísérletben résztvevő hallgatókat leszíjjazták, extrém stressznek tették ki, miközben reakcióikat elektródákkal mérték, dühreakcióikat többször visszajátszották nekik, folyamatosan megszégyenítették őket. A legtöbb kutatóintézet és egyetem etikai kódexe igencsak szigorú manapság. A kutatás alanya csakis önkéntes alapon vehet részt egy kutatásban, tudnia kell arról, hogy milyen kutatás része, és ehhez aktivan hozzá is kell járulnia, valamint joga van anonim maradni. A kutató köteles garantálni, hogy a kutatásban résztvevők sem fizikailag sem lelkileg nem károsodnak amiatt, hogy részt vesznek a kutatásban, az adataikat a GDPR szabályoknak megfeleően kell kezelni, valamint csak speciális etikai engedéllyel alkalmazhat a kutató olyan résztvevőket, akik valamilyen oknál fogva alárandelt helyzetben vannak.
A Stanford- , a Milgram-, avagy a Tuskagee szifilisz kisérletet ma biztosan nem engedélyezné egyik egyetem sem, mint ahogy amit Dr Murray művelt 1960-ban a fiatal Theodore Kaczynskivel, akiből a hírhedt Unabomber lett, ma már megengedhetetlen: a kísérletben résztvevő hallgatókat leszíjjazták, extrém stressznek tették ki, miközben reakcióikat elektródákkal mérték, dühreakcióikat többször visszajátszották nekik, folyamatosan megszégyenítették őket. A legtöbb kutatóintézet és egyetem etikai kódexe igencsak szigorú manapság. A kutatás alanya csakis önkéntes alapon vehet részt egy kutatásban, tudnia kell arról, hogy milyen kutatás része, és ehhez aktivan hozzá is kell járulnia, valamint joga van anonim maradni. A kutató köteles garantálni, hogy a kutatásban résztvevők sem fizikailag sem lelkileg nem károsodnak amiatt, hogy részt vesznek a kutatásban, az adataikat a GDPR szabályoknak megfeleően kell kezelni, valamint csak speciális etikai engedéllyel alkalmazhat a kutató olyan résztvevőket, akik valamilyen oknál fogva alárandelt helyzetben vannak.
Sam Kean 2021-es könyve jócskán belenyúl az erkölcstelen, törvénytelen és gonosz tudomány témakörébe, és a klasszikus etikailag igencsak megkérdejelezhető esetek mellett hoz különös és meghökkentő példákat. A jégcsákányos műtős, Walter Freeman, aki egy jécsákánnyal hajtott végre az orron keresztül lobotómiát Amerika-szerte mentális betegeken és akiről a könyv a címét is kapta, különösen megdöbbentő és brutális, akárcsak Edison villamosszékes állatkísérletei. A kalóz biológus és a rabszolgakereskedő természetbúvár története az enyhébb és talán az emberileg érthetőbb sztorik közé tartoznak, bár az általuk feltett erkölcsi kérdések nem kevésbé súlyosak. Aztán vannak persze horror történetek is, mint például az emberölésre átnyergelő hullarablók esete, akik az orvosképzés boncolásaihoz szállították a holttesteket, avagy az áldozatát feldaraboló vegyészprofesszor esete.
Kémkedés, tudományos államtitkok ellenséges országoknak való eladása, dinócsontok körüli elmérgesedett vita, csalás, van itt (és ez alatt a tudományos kutatás és orvostudomány értendő) kérem minden, amit csak egy szaftos bulvárlap kivánhat.
Hogy mért is érdemes elolvasni ezt a könyvet? Kean mesterien fűzi fel a hátborzongató történetek egy szálra: a tudomány nem választható el a társadalomtól. Együtt éljük hétköznapjainkat mi kutatók, orvosok, tudósok a hétköznapi emberekkel, és a tudomány eredményi hatással vannak mindnyájunkra. A tudomány, mint fogalom, nem lehet feljebbvalóbb az emberi életnél, és az emberi értékrendet be kell épiteni a tudományos kérdések megvitatásába.
Víg Julianna
 Minden út a sejtbiológiához vezet – ha egy tőmondatban kellene összefoglalnom Siddhartha Mukherjee legújabb könyvének tanulságát, valószínűleg ennyit mondanék. Talán azért is tetszett annyira a könyv, mert felidézte, mennyire fel voltam villanyozva, amikor először vontam le ezt a tanulságot. Ez még boldog biológushallgató koromban történt, amikor már tanultam annyit az élőlényeknek mind a sejt “alatti” szerveződési szintjéről, vagyis a molekulák és anyagcserefolyamatok világáról, mind a sejt “felettiről”, vagyis az anatómiáról és élettanról, hogy észrevegyem: a sejtbiológiában ez a kétféle lépték és tudás találkozik. Pontosan ezt a csomópont-jelleget emeli ki Mukherjee is, aki egy-egy sejttípuson, illetve sejtfunkción keresztül mutat be egymástól – látszólag – igen távoli jelenségeket, a mesterséges megtermékenyítéstől az őssejtterápián, a rákon és a “fiatalító” vérátömlesztésen át a COVIDig.
Minden út a sejtbiológiához vezet – ha egy tőmondatban kellene összefoglalnom Siddhartha Mukherjee legújabb könyvének tanulságát, valószínűleg ennyit mondanék. Talán azért is tetszett annyira a könyv, mert felidézte, mennyire fel voltam villanyozva, amikor először vontam le ezt a tanulságot. Ez még boldog biológushallgató koromban történt, amikor már tanultam annyit az élőlényeknek mind a sejt “alatti” szerveződési szintjéről, vagyis a molekulák és anyagcserefolyamatok világáról, mind a sejt “felettiről”, vagyis az anatómiáról és élettanról, hogy észrevegyem: a sejtbiológiában ez a kétféle lépték és tudás találkozik. Pontosan ezt a csomópont-jelleget emeli ki Mukherjee is, aki egy-egy sejttípuson, illetve sejtfunkción keresztül mutat be egymástól – látszólag – igen távoli jelenségeket, a mesterséges megtermékenyítéstől az őssejtterápián, a rákon és a “fiatalító” vérátömlesztésen át a COVIDig.
A biológiai-orvosi fogalmakat, folyamatokat Mukherjee ezúttal is mindenki számára érthető és – mint minden nagy ismeretterjesztő – sokszor egészen költői metaforákkal írja le. Emellett fontos alkotóeleme a könyvnek a sok (fotókkal is illusztrált) tudománytörténeti érdekesség, amelyek közt értelemszerűen kiemelt szerepet kapott a sejtelmélet megszületése, és ezen túl a mikrobiológia, az immunológia, a genetika és az idegtudomány néhány megunhatatlan sztorija. A sztorik közt két magyar vonatkozású is van: Semmelweis Ignác mellett Nagy László szegedi evolúcióbiológus nevét is kiszúrhatja a figyelmes olvasó.
A tudomány és a tudománytörténet mellé igen sok személyes jellegű történetet is kapunk. Bevallom, nekem helyenként túl sok, túl amerikai a személyesség és a dráma. Mindjárt a könyv első mondata arról szól, hogy a szerző végignézte egy barátja halálát, de torokszorító részleteket olvashatunk a halál küszöbéről visszahozott kislányról, és a szerző saját visszatérő depressziójáról is. Persze ha belegondolok, hogy Mukherjee karrierjének jelentős részét rákbetegek gyógyítására tette fel, mindjárt megértőbb vagyok a drámai tónussal. (Legalább a könyveiben hadd drámázzon egy kicsit.) És abban viszont biztos vagyok, hogy a sejtek énekébe belehallgatni nemcsak egy nehéz szívű onkológus számára segíthet kizárni a nyomasztó gondolatokat, hanem bármelyikünknek, aki menekülne néha a szabad szemmel látható világ elől.
Zsámboki János
Kevés olyan pálfordulás ismert a zöld mozgalom történetében, mint Mark Lynasé. Ő ugyanis vagy húsz éven át GMO ellenes aktivistaként működött, a kilencvenes évektől kezdve. Azonban mindenki meglepetésére 2013 -ban nyilvánosan ismerte el, hogy mekkorát tévedett a kérdésben, ennek az előadásnak a felvétele azóta is népszerű. Néhány évvel később, 2018-ban jelent meg a könyve, a „Seeds of Science„, amit részben magyarázatnak szánt, részben önfelmentésnek, részben útmutatásnak.
Az első fejezetek a kilencvenes évekbe repítenek vissza, a mozgalmi időket idézi meg, leírja, hogyan került a GMO ellenes mozgalomba, milyen akciókat hajtottak végre. Nagyon szemléletes, hogy leírja, az aktivisták közül soha senkinek semmilyen valódi ismerete sem volt a technológiáról, az egész nagycégezést, a GMO ellenességet tokkal-vonóval átvették a tengerentúlról. Bár a GMO ellenességet mindig is alulról építkező népmozgalomként akarták eladni, már a kilencvenes években is egy az egyben életvitelszerű aktivisták reklámkampánya volt.A második fejezet arról szól, hogyan gondolta meg magát Mark. Meglepő módon felhívták a Guardian újságtól, hogy valamilyen miniszter nem átallott pozitívan nyilatkozni a GMO -król, úgyhogy kéne valami ellenmaszlag, ő pedig készségesen meg is írta egy óra alatt, hiszen „csukott szemmel is meg tudott már ekkor írni egy gyors anti-GMO szöveget”. Ez után teljesen meglepődött, hogy az újságban megjelent írására a válaszok nagyrészt negatívak, konkrétan ki is mondták, hogy mélyen tudományellenes a szösszenete, a GMO ellenesség Európa kreacionizmusa és hasonlók. Ekkor már komoly tudományos ismeretterjesztő munkásság állt a háta mögött a klímaváltozásról, föl is háborodott rajta, hogy miért nem hiszik el neki a szokásos maszlagot, bár ugye azt el kellett ismernie, hogy nem sok tudományos bizonyítékot mellékelt. Úgyhogy nekiállt összeszedni a bizonyítékokat, hogy ne kritizálhassák azzal, hogy tudományellenes a mondókája. De itt kezdődött a törés, ugyanis bizony a szakirodalomban nem talált semmi olyasmit sem, amit addig állított. Oké, akkor vissza akart nyúlni a szokásos módszerhez és idézni akarta a nagyobb tekintélyű tudományos testületek állásfoglalásait, amik alátámasztanák az állításait. De ezeket sem találta. Ekkor kezdett rájönni, hogy ebben a kérdésben ő, a zöld mozgalom és a teljes liberális társadalom éppen a tudományellenes oldalon áll.
A könyv hátralévő része a valóságot próbálja körüljárni. A harmadik fejezet a rekombináns DNS technológia történetét meséli el, a negyedik a Monsantóét.
 Az igazán érdekes részek viszont az ötödik fejezettől kezdődnek. Ugyanis Mark Lynas nem átall utánamenni a zöld mozgalom tevékenységének az ügyben. Körbejárja Vandana Shiva hazugságait az indiai farmeröngyilkosságokról, érdekes elolvasni, hogyan sikerült a zöld mozgalomnak egy teljesen hazug történetet eladni a teljes nyugati sajtónak amit gyakorlatilag a mai napig alapvetően igaznak tart mindenki. Ez a hazug valóságteremtés az egész könyvön végigvonul, ahol megjelent a zöld mozgalom, ott bizony azonnal terjedni kezdtek a nevetségesen valótlan történetek. Egészen félelmetes például, hogy a bangladesi Bt-toxin termelő padlizsán kipróbálásakor egészen szervezett mozgalmat találtak, akik személyesen keresték fel a programban résztvevő parasztokat, hogy lejárató hazugságokkal tömje a fejüket a GM-padlizsánról. Például írt egyszer egy Mohammed Hafizur Rahman nevű bangladesi parasztról, aki nagyon elégedett volt a GM-padlizsánnal, amit termesztett. Miután a történet megjelent a New York Timesban, bár szándékosan nem írta le, hol is él Mohammed, azért az aktivisták csak megtalálták a százhatvan millió lakosú Bangladesben és rögtön elkezdték agitálni is, hogy valójában milyen szörnyű dolog a GM-padlizsán, nehogy szegény a saját szemének higgyen.
Az igazán érdekes részek viszont az ötödik fejezettől kezdődnek. Ugyanis Mark Lynas nem átall utánamenni a zöld mozgalom tevékenységének az ügyben. Körbejárja Vandana Shiva hazugságait az indiai farmeröngyilkosságokról, érdekes elolvasni, hogyan sikerült a zöld mozgalomnak egy teljesen hazug történetet eladni a teljes nyugati sajtónak amit gyakorlatilag a mai napig alapvetően igaznak tart mindenki. Ez a hazug valóságteremtés az egész könyvön végigvonul, ahol megjelent a zöld mozgalom, ott bizony azonnal terjedni kezdtek a nevetségesen valótlan történetek. Egészen félelmetes például, hogy a bangladesi Bt-toxin termelő padlizsán kipróbálásakor egészen szervezett mozgalmat találtak, akik személyesen keresték fel a programban résztvevő parasztokat, hogy lejárató hazugságokkal tömje a fejüket a GM-padlizsánról. Például írt egyszer egy Mohammed Hafizur Rahman nevű bangladesi parasztról, aki nagyon elégedett volt a GM-padlizsánnal, amit termesztett. Miután a történet megjelent a New York Timesban, bár szándékosan nem írta le, hol is él Mohammed, azért az aktivisták csak megtalálták a százhatvan millió lakosú Bangladesben és rögtön elkezdték agitálni is, hogy valójában milyen szörnyű dolog a GM-padlizsán, nehogy szegény a saját szemének higgyen.
A hatodik fejezet Afrikáról szól, ahol egy az egyben sikerült megfojtani a GM-növényeket, kategorikusan betiltották mindet a legtöbb államban. A hetediktől a tizedik fejezetig a könyv a GMO ellenes mozgalommal foglalkozik. Érdekes elolvasni, hogyan indult a mozgalom, hogyan szerveződik. A legérdekesebb viszont a „Hogyan gondolkodnak a környezetvédők?” fejezet. Érdemes elolvasni, mivel éppen azt próbálja meg feltárni, hogyan lehetne hatni erre a mozgalomra, hogyan lehetne meggyőzni őket.
Kun Ádám
Dunbar azon kevesek egyike, akiről egy számot elneveztek. Ez a Dunbar-szám megmondja, hogy mi a korlátja az ismerőseink számának, ami 150. Persze ennél több embernek a nevére emlékezhetünk, facebookon lehet tízszer ennyi „barátunk”, de attól még minden kutatás azt mutatja, hogy ennyi emberrel vagyunk ténylegesen kapcsolatban. Barátokra pedig szükségünk van, mert csoportos élőlényként elég rosszul működünk társak és társas kapcsolatok nélkül.
 A könyv a társas kapcsolatok előnyeivel indít és a kapcsolati hálózat feltérképezésével folytatódik. Egy nagyon szűk időablak állt a kutatók rendelkezésére, amikor már volt elég digitális adat a telefonhívások, mobil cellaadatok, iwiw és korai facebook (és társai) platformok jóvoltából, de még nem állt rendelkezésre túl sok ilyen, hogy még lehessen automatikusan (de anoním módon) követni, hogy ki kivel áll kapcsolatban. Ma már szinte mindenkivel más platformon kommunikálunk, így mindegyik hamis képest mutat. Marad a kérdőívezés, ami viszont egyre nehezebb, ahogy az emberek egyre kevésbé szeretnék, ha túl sok adat gyűlne róluk.
A könyv a társas kapcsolatok előnyeivel indít és a kapcsolati hálózat feltérképezésével folytatódik. Egy nagyon szűk időablak állt a kutatók rendelkezésére, amikor már volt elég digitális adat a telefonhívások, mobil cellaadatok, iwiw és korai facebook (és társai) platformok jóvoltából, de még nem állt rendelkezésre túl sok ilyen, hogy még lehessen automatikusan (de anoním módon) követni, hogy ki kivel áll kapcsolatban. Ma már szinte mindenkivel más platformon kommunikálunk, így mindegyik hamis képest mutat. Marad a kérdőívezés, ami viszont egyre nehezebb, ahogy az emberek egyre kevésbé szeretnék, ha túl sok adat gyűlne róluk.
A barátok, ismerősök és kapcsolati hálózatok az egyénektől függenek. Az egyének neurális és hormonfolyamatai befolyásolják a viselkedésüket, de a viselkedés befolyásolja a barátokat, akikkel való viszonyunk kihat a mentális állapotunkra és a hormonjainkra. Tehát a barátságok hatását a szervezeten belül is lehet vizsgálni. De a barátok, ismerősök és kapcsolati hálózatok alkotják a társadalom szövetét, így egy az egyénnél sokkal magasabb szinten is vizsgálhatjuk hatásait. A könyv természetesen az együttműködésre is kitér. Miben mások a közösségeink egy csimpánz közösséghez képest? Hogyan járul hozzá ez ahhoz, hogy ki tudtunk alakítani technikai civilizációt, nagyfokú együttműködést?
Dunbar munkásságának egy részét ismertem, így azok visszaköszönnek a könyv lapjairól. Persze sokkal emészthetőbb formában, mint a szakcikkekből. És persze felhívja a figyelmemet, hogy mit nem olvastam még, pedig kellett volna. A nyelv evolúciójáról még vannak fura elképzelései, de ez bocsánatos bűn, nincs vele egyedül.
Kun Ádám
 Ez a könyv rólunk szól, ahogy szinte minden a biológiában. Ez a könyv az érzékelésünkről szól. A legtöbb példa mégsem az emberről szól, bár nagyobb részében olyan érzékekről, amelyekkel mi is rendelkezünk és csak a végén megemlítve párat, amivel egyáltalán nem. Ennek ellenére ez a könyv azt mutatja meg, hogy mi milyenek vagyunk azon keresztül, hogy milyenek és mennyire mások a körülöttünk levő élőlények. Továbbá végigvonul a könyvön az a gondolat, hogy a saját magunk érzékei által korlátozott világ egyben mennyire nehézzé teszi más élőlények megértését.
Ez a könyv rólunk szól, ahogy szinte minden a biológiában. Ez a könyv az érzékelésünkről szól. A legtöbb példa mégsem az emberről szól, bár nagyobb részében olyan érzékekről, amelyekkel mi is rendelkezünk és csak a végén megemlítve párat, amivel egyáltalán nem. Ennek ellenére ez a könyv azt mutatja meg, hogy mi milyenek vagyunk azon keresztül, hogy milyenek és mennyire mások a körülöttünk levő élőlények. Továbbá végigvonul a könyvön az a gondolat, hogy a saját magunk érzékei által korlátozott világ egyben mennyire nehézzé teszi más élőlények megértését.
Engem kimondottan zavar a sötétség, főleg, ha kint az utcán szeretnék járni. Ezért van közvilágítás a településeinken. A nappali életmódú emberszabású nappallá (na jó, szürkületté) változtatja az éjszakát maga körül. És közben nem gondolunk bele, hogy ez mennyire megzavar minden más élőlényt, amelyek evolúciósan megszokták, hogy van világos és van sötét időszak és nem világos és kevésbé világos időszak váltogatja egymást. Rotorhanggal zajongjuk tele az óceánokat, tükröződő felületeket rakunk oda, ahol olyan sohasem volt, és új szagokkal terheljük a levegőt. A rotor mély hangját alig halljuk, az ablakok nekünk üvegfelületnek látszanak és nem víznek, a szagokkal meg – a vasárnapi rántotthúson és egymás szagán kívül – nem nagyon törődünk (pedig érzékelni képesek vagyunk azokat). Ha ennyire nem tudunk mit kezdeni a kicsit más érzékeléssel, akkor mégis hogyan képzeljük el a világot a színek egy teljesen új dimenziójával, amivel igen sok nem-emlős rendelkezik? Vagy milyen lehet lényegében panorámában látni a világot, mint több növényevő? Esetleg a kezünkkel ízlelni, hogy már akkor tudjuk, hogy mennyire ehető valami, amikor hozzáértünk?
A könyv hosszú. Rengeteg muníció van benne egy baráti beszélgetés feldobására. Tudod, hogy a harcsák egész bőrfelületén vannak ízlelőbimbók? Mintha egy óriás nyelv lenne az egész állat. Tudod, hogy egyes rovarok „füle” a lábán van, vagy a szárnya tövében? A könyv nagy része ilyenekkel van tele, és azzal, hogy a kutatók hogyan jöttek rá, hogy mit érzékelnek az állatok. Már, amely állatok esetében tudjuk.
Ami kevésbé tetszett a könyvben az a nekem erőltetett interjúérzet, hogy mindent egy-egy amerikai kutató szemszögéből kell látnunk. Mások eredményei pedig lábjegyzetbe kerülnek, pedig megérdemelnék a főszöveget is. Lábjegyzetből sok van, túl sok.
Nekem ez az első Ed Yong könyvem. Itt is méltatták már és nem kellett csalódnom benne. Tényleg jól ír.
Varga Máté
Ritkán fordul elő, bár nem példátlan, hogy ketten is írunk ugyanarról a könyvről, de Ed Yong könyve egyébként is megérdemel egy dupla-kritikát. Bár már blogger korában is szerettem Yongot és az általa még 2006-ban, valamikor a természettudományos blogírás fénykorában elkezdett Not Exactly Rocket Science az egyik kedvenc hírforrásom volt anno, igazából azt hiszem nem vitatható, hogy jelenlegi munkahelyén a The Atlantic-nél vált belőle korunk egyik legjobb ismeretterjesztője, nem véletlen a 2021-es Pulitzer díj. (BTW, a CriticalBiomass de facto néhány hónappal korábban indult és még hivatalosan nem zártuk le, szóval ez is egyfajta apró győzelem ;-).)
Már a Pulitzert megelőző „I contain multitudes” is minden idők top ismeretterjesztője közé emelte Yongot, nem véletlenül ajnároztam már 2017-ben is, ez a műve azonban – szerintem – még azon is túltesz.
Bizonyos szempontból tökéletes párja ez a könyv a tavaly bemutatott, Peter Godfrey-Smith jegyezte Metazoának, csak míg utóbbi kifejezetten azt feszegette, hogy evolúciósan miképp alakultak ki az egyre bonyolultabb felépítésű idegrendszerek és ez milyen új kognitív működést tett lehetővé egyes állatcsoportokban, Yong könyve azt mutatja be szemléletesen, hogyan, hány féleképpen látják világunkat közeli és távoli rokonaink ezen a bolygón.
A könyv kulcsszava az Umwelt, a Jakob Johann von Uexküll által bevezetett kifejezés, ami körülbelül azzal egyenrétékű, hogy egy állatnak, saját érzékszervei felhasználásával, milyen szubjektív érzete lehet a külvilágról. És, ahogy Ádám is utalt rá, bizony újból és újból fény derül arra, hogy az emberi Umwelt is szegényes, sok dolgot el sem tudunk képzelni. Mert lehetnek hasonlataink arra, hogy milyen lehet, amikor egy csörgőkígyó hőreceptorokkal is látja áldozatát, vagy egy denevér echolokációval közelíti meg az áldozatául szolgáló lepkét, de ezek minden jel szerint szegényes próbálkozások annak a szenzoros érzetnek a leírására, amit megfelelő receptorok hiányában istenigazából igazából nem tudunk elképzelni és felfogni.
Azt, ha egy-egy faj más hullámhosszokon (is) látja a világot, mint még csak-csak felfogjuk, ahogy azt is, hogy egyes rovarfajok ízérzékelés és akár hallása a lábukon keresztül történik, hiszen hasonló ingereket feldolgozó receptoraink nekünk is vannak. De hogy lehet elképzelni azt, hogy egy delfin érzékeli a környezetében levő tárgyak és lények sűrűségét, vagy, hogy egy teknős a bolygó mágneses terét is használja a tájékozódáshoz? Bármennyire is sok dolgot el tudunk képzelni, ilyen érzékelésre képtelenek vagyunk, a világ bizonyos szegletei számunkra sötétben vannak és maradnak, és szenzoros plátói barlangunkban ilyen esetekben szegényesnek bizonyuló hasonlataink adnak bármi információt ezeknek a fajoknak a valóságérzetéről.
Varga Máté
Szomorú apropója miatt, talán ez volt a tavalyi év legfontosabb tudományos ismertető könyve. Quammen nem kevesebbre vállalkozott, mint, hogy megírja a COVID-19 járvány lehető legpontosabb történetét: hogy fedezték fel, hogyan terjedt el, honnan eredhetett és hogyan evolválódott.
 Mivel a fent említett témákról minden bizonnyal karaktermilliókat írtak az elmúlt évek során (csak az előbb említett Ed Yong is több könyvnyi hosszúságú cikket írt a The Atlantic-be erről a témáról), mindenkiben felmerülhet, hogy van-e bármi szükség egy ilyen könyvre. Az én véleményem nem csak az, hogy igen, de egyben ez az a könyv, amire konkrétan szükség van.
Mivel a fent említett témákról minden bizonnyal karaktermilliókat írtak az elmúlt évek során (csak az előbb említett Ed Yong is több könyvnyi hosszúságú cikket írt a The Atlantic-be erről a témáról), mindenkiben felmerülhet, hogy van-e bármi szükség egy ilyen könyvre. Az én véleményem nem csak az, hogy igen, de egyben ez az a könyv, amire konkrétan szükség van.
Quammen eszméletlen mennyiségű szakirodalmat dolgoz fel, ráadásul személyes interjúkban hihetetlen mennyiségben és minőségben dokumentálta, mi történt. A járvány elején, szinte órákra lebontva követhetjük, hogy próbálják az első vírusgenomot megszekvenálni, összeszerelni és nyilvánosságra hozni ausztrál, kínai és amerikai kutatók. Később nagyobb időléptékre váltunk, de nem kevésbé olvasmányosak azok a részek, ahol a vírus-evolúciót célzó kutatásokba láthatunk bele, amelyek a dominánsá váló variánsok kiszúrását tűzték célul, vagy hogy miként tudott a SARS-CoV-2 vírus futótűzszerűen elterjedni a minket körülvevő állatokban is.
Ez utóbbi átköt abba a járvány eredetét firtató, sokakat lázba hozó kérdésekbe is. Ugyan Quammen végig hangsúlyozza, miért és mennyire nehéz ezekre választ adni (a fejezet címe is ezt tükrözi: „Nobody Knows Everything„) és bőven teret ad olyanoknak is, mint Alina Chan, akik konzekvensen a járvány „laboreredete” mellett törtek lándzsát, aligha marad kétséges a fejezet olvasása után, hogy a szerző messze valószínűbbnek tartja, hogy az egész egy banális zoonózissal kezdődött a Huanan piacon.
Varga Máté
 Balcombe könyvéről nem hallottam korábban, de amint megláttam, tudtam, hogy el kell olvasnom: amolyan szakmai kötelesség volt, hiszen saját csoportom kísérleteinek egy része is különböző halfajok kognitív képességeivel foglalkozik.
Balcombe könyvéről nem hallottam korábban, de amint megláttam, tudtam, hogy el kell olvasnom: amolyan szakmai kötelesség volt, hiszen saját csoportom kísérleteinek egy része is különböző halfajok kognitív képességeivel foglalkozik.
A könyv címe (bennem) megidézte a legendás „A belső hal (Your Inner Fish)” című könyvet, ahol Neil Shubin anatómiai és fejlődéstani hasonlóságokon keresztül mutatja be, mi minden közös a gerincesekben (vagyis akár bennünk és egy halban is). Abból a szempontból helytálló a hasonlat, hogy Balcombe tényleg hosszan mutogat olyan tulajdonságokat, amelyeket a legtöbb laikus emlős-, vagy akár ember-specifikusnak gondolhat, pedig nagyon nem az és „vízben élő unokatestvéreink” esetében is megfigyelhető.
Messze nem triviális, hogy honnan tudja egy géb, miután párszor elúszott egy rögös felszín fölött dagálykor, hogy apálykor milyen irányba és mekkorát kell ugrania, hogy egyik visszamaradt pocsolyából pontosan szomszédosba kerüljön és ne a partra vetett hal effektust mutassa be. Hogyan találnak vissza több ezer kilométeres vándorlás után lazacok a szülőhelyükre? De ugyanígy nem triviális, honnan ismerik fel a tengeri tisztogató halak, melyik hal úszkál körülöttük kuncsaftként és melyik potenciális predátorként (akár ugyanabba a fajba tartozó egyedek közül) és hogyan tartják fejben, akár egyedi szinten is, ki a jó kuncsaft és ki nem az, kivel érdemes kivételezni és kivel nem.
Mondhatjuk-e egy halról, hogy játszik, ha láthatóan céltalan tevékenységet végez, ami nem emlékeztet olyan veleszületett mozgáskombinációkra, ami például a táplálkozáshoz fontos. Arról már nem is szólva, hogy különböző halfajokban milyen széles tárháza jelenik meg a szülői gondoskodásnak: nem minden faj hagyja ott az utódait az ívás után, van aki fészket épít, van aki szájában hordozza őket.
A könyv előnye, hogy nagyon széles hálóval merít az anekdotikus esetekből, s ez egyben a hátránya is: számos olyan jelenséget ír le a szerző, ahol messze nincs bizonyítva, hogy az az adott fajra jellemző tulajdonság, sőt, néha még a megfigyelések megismételhetősége is kérdéses. Mivel ez nem tankönyv, hanem szórakoztató ismeretterjesztés, ez önmagában nem gond, viszont amikor figyelembe vesszük Balcombe tágabb (de nem titkolt) célját, akkor már problémássá válik: már a könyv előszavából kiderül, hogy a szerző azt szeretné bebizonyítani nekünk, hogy a halak ugyanolyan érző lények, mint mi, így nem csak a túlhalászás és embertelen tömegtartás a problémás, hanem tulajdonképpen szinte bármilyen tevékenység, aminek halfogyasztás a vége (legyen az akár mezei horgászat), hiszen érző, gondolkodó lényekről van szó. Ami ebbe a világképbe már kevésbé fér bele és ennek megfelelően nem is nagyon kap helyet a könyvben, hogy a halfajok többsége lelkiismeretlen predátor is egyben és minden skrupulus nélkül elfogyasztanak más halakat, legyenek azok fajtársak, vagy akár saját utódaik…
Varga Máté
Humán genetikai és populációgenetikai körökben ez volt talán a közelmúlt legviharosabban fogadott könyve és a ha csak egyes kritikákat olvasnánk, vagy az azok nyomán is kirobbanó Twitter-vitákat, könnyen juthatnánk arra következtetésre, hogy Harden egy titkos, vagy nem is olyan titkos eugenista célzattal írta ezt a könyvet.
„GWAS undoubtedly captures some causal genetic effects, i.e., more than confounding alone, and there is interesting science to learn from these initial findings.” pic.twitter.com/HOQgqV1OXP
— Kathryn Paige Harden (@kph3k) January 25, 2022
A viharos fogadtatás legfőbb oka értelemszerűen a kényes téma: mennyire felelősek génjeink összetett tulajdonságainkért, mennyire van a génjeinkbe írva, hogy mi lesz a végzettségünk, vagy pont, hogy mennyi lesz a keresetünk. A 20. századi eugenikai „megoldások” érthető okokból mély nyomokat hagytak sokakban és hosszú időre tabuvá tették annak a gondolatát is, hogy komplex tulajdonságoknak, viselkedésformáknak genetikai alapjuk lehet. Ezért is sokak által osztott álláspont, amely szimplán tagadja, hogy a genetika és az emberi viselkedés között kapcsolat lehet és minden viselkedés esetében kizárólagosan tanult magatartásformákat keres. Extrémebb esetekben ez oda vezethet, hogy már minden olyan kérdésfeltétel, ami ezt nem eleve axiómaként fogadja el, gyanús és hátsó szándékot sejtet.
 Harden egész karrierje márpedig arról szól, hogy ezt, a szerinte dogmatikus álláspontot megkérdőjelezze. Talán a legfontosabb „eszköze” ebben a munkában az ún. GWAS-analízis, amely segítségével, genom-szinten lehet egyes esetekben olyan polimorfizmusokat találni, amelyek bizonyos jellegekkel együtt jelennek meg. Ezek a GWAS tanulmányok voltak az elsők, amelyek elkezdték pedzegetni, hogy bizonyos szociokulturális jelenségeknek genetikai hátterük lehet (vagyis lehet olyan bonyolult jelleget találni, ami mögött genetikai okok vannak), de mivel őrülten nehéz megfelelő kontrollokat alkalmazni ezekben a tanulmányokban, plusz eleve több százezres mintaszám kell ahhoz, hogy egyáltalán komolyan vehető eredmények születhessenek, a genetikusok jelentős része még ma is sokszor kétségbe vonja az ilyen típusú eredményeket. (Graham Coop bevezetőjét mindenképpen ajánlom a téma iránt érdeklődőknek.)
Harden egész karrierje márpedig arról szól, hogy ezt, a szerinte dogmatikus álláspontot megkérdőjelezze. Talán a legfontosabb „eszköze” ebben a munkában az ún. GWAS-analízis, amely segítségével, genom-szinten lehet egyes esetekben olyan polimorfizmusokat találni, amelyek bizonyos jellegekkel együtt jelennek meg. Ezek a GWAS tanulmányok voltak az elsők, amelyek elkezdték pedzegetni, hogy bizonyos szociokulturális jelenségeknek genetikai hátterük lehet (vagyis lehet olyan bonyolult jelleget találni, ami mögött genetikai okok vannak), de mivel őrülten nehéz megfelelő kontrollokat alkalmazni ezekben a tanulmányokban, plusz eleve több százezres mintaszám kell ahhoz, hogy egyáltalán komolyan vehető eredmények születhessenek, a genetikusok jelentős része még ma is sokszor kétségbe vonja az ilyen típusú eredményeket. (Graham Coop bevezetőjét mindenképpen ajánlom a téma iránt érdeklődőknek.)
Harden, aki egyébként baloldalinak vallja magát, nem azért foglalkozik mindezzel (mint azt kritikusai néha sarkítva állítják), hogy bebizonyítsa, vannak olyan emberek, akik „genetikailag predesztináltak” arra, hogy csak érettségijük legyen és azokkal nincs mit tenni, kár pénzt fordítani az oktatásukra (ez a klasszikus eugenista nézet, napjainkban altright topszként szokott fel-feltűnni). Az ő érvelése pont fordított: ha valóban léteznek olyan faktorok, amelyek a mai rendszerben sok embert genetikailag predesztinálnak arra, hogy rosszabbul teljesítsenek, akkor fontos kutatnunk őket, mert csak akkor tudunk társadalmi szinten javítani a helyzetükön. Egyszerű hasonlatban, gondoljunk arra, hogy a genetikai okokból kialakuló rövidlátást kezelhetjük úgy is, hogy a rövidlátóknak nem segítünk, sőt egyre távolabb ültetjük őket a táblától, megideologizálva azzal, hogy a jól látóktól veszik el a jobb helyeket, pedig nekik amúgy is mindegy, vagy úgy is, hogy szemüveget adunk nekik, ezzel kiegyenlítve genetikai hátrányukat.
Harden gondolatai nagyon is relevánsak napjaink géndiagnosztikáról és génszerkesztésről szóló vitáiban és a könyv sokat tesz azért, hogy ezt az egyébként nem könnyű (de sokakat érdeklő) témát könnyebben emészthetővé tegye.
(Akit kicsit bővebben is érdekel a téma, annak ajánlom Harden és Carl Zimmer beszélgetését a könyv apropóján.)
Varga Máté
Látszólag kilóg az aktuális könyvek ajánlójából ez a mű, hiszen finoman szólva sem aktuális könyvről van, szó, de tavaly is előhozakodtam egy hasonló kaliberű (és szintén nem épp friss) Richard Lewontin művel, ezt a „hagyományt” folytatnám ezzel. Dobzhansky szintén a múlt századi biológia legendája, akik oroszlánrészt vállalt a korabeli genetika és evolúcióbiológia téziseinek összeboronálásában, az ún. „új szintézis” megalkotásában.
 Gyerekkoromban, a nappalinak egy jól látható helyén, színes, kisméretű könyvekből álló sorozat hívta fel magára a figyelmet. Engem (akkor) a témák nem nagyon fogtak meg – kellett bő évtized, hogy ez megváltozzon – de mindenképpen látszott, hogy a Kriterion Kiadó Téka sorozata nagyon széles témakört ölel fel: megfért benne Tacitus és Francis Bacon, Moholy-Nagy László és Marcus Aurelius. Ma már persze rácsodálkozom, hogy a szerkesztők milyen fantasztikus érzékkel válogattak össze ma is értékes és érdekes műveket, ahol ókori filozófusoktól egészen a modern tudományig minden képviseltette magát. Persze a szerkesztői előszavakat érdemes volt akkor is átugorni, hiszen azok tulajdonképpen nem az olvasónak, hanem a ceaușescui cenzoroknak szóltak (elmagyarázva, hogy a dolgozó román népnek miért hasznos az adott mű kiadása), de ezen túl kifejezetten színes világok keltek a lapokon életre.
Gyerekkoromban, a nappalinak egy jól látható helyén, színes, kisméretű könyvekből álló sorozat hívta fel magára a figyelmet. Engem (akkor) a témák nem nagyon fogtak meg – kellett bő évtized, hogy ez megváltozzon – de mindenképpen látszott, hogy a Kriterion Kiadó Téka sorozata nagyon széles témakört ölel fel: megfért benne Tacitus és Francis Bacon, Moholy-Nagy László és Marcus Aurelius. Ma már persze rácsodálkozom, hogy a szerkesztők milyen fantasztikus érzékkel válogattak össze ma is értékes és érdekes műveket, ahol ókori filozófusoktól egészen a modern tudományig minden képviseltette magát. Persze a szerkesztői előszavakat érdemes volt akkor is átugorni, hiszen azok tulajdonképpen nem az olvasónak, hanem a ceaușescui cenzoroknak szóltak (elmagyarázva, hogy a dolgozó román népnek miért hasznos az adott mű kiadása), de ezen túl kifejezetten színes világok keltek a lapokon életre.
A Korunk áprilisi számába írtam a muslicák tudománytörténetéről, s ez adta az apropóját, hogy végre elolvassam Dobzhansky 1973-as „Genetic Diversity and Human Equality” című könyvének 1985-ös Tékás fordítását (egyébként, ha jól tudom, más fordítása nem is létezik ennek a könyvnek). Dobzhansky maga az egyik legfontosabb alakja volt a múlt század második felében fellángoló „nature” vs. „nurture” vitának, ahol a saját kísérleti tapasztalait felhasználva érvelt szenvedélyesen a mellett, hogy a társadalom sokszínűsége (genetikai és nem csak) egy természetes és hasznos dolog és mindaz az eugenikai irányvonal, amelyet például a Dobzhansky-hoz hasonlóan Thomas Hunt Morgan laborjából kikerült Herman Joseph Muller hirdetett, nem csak hamis, de veszélyes is. A Mullerhez hasonló „genetikai Jeremiások”, akik folyton az emberi genetikai állomány devalvációjától tartottak és pozitív genetikai programok (pl. mesterséges megtermékenyítés Nobel díjas férfiak spermájával) keretében akarták ezt az imminens armageddont elkerülni, nem értették, hogy miként működik az evolúció, hogyan keveri át a genetikai anyagot és miként válhat a sokszínűség a pont a populáció előnyévé új szelekciós nyomás alatt. Mindez nem azt jelentette, hogy Dobzhansky elvetette volna a genetika szerepét akár az összetett jellegek kialakulásában. Ellenkezőleg, de az ebből adódó társadalmi problémákra szerinte nem az eugenika korai, vulgáris formája adhatta meg a választ. Ilyen szempontból, talán azt is mondhatjuk, hogy a korábban emlegetett Paige Harden előfutára mindaz, amit Dobzhansky ebben a művében is papírra vetett.
Nem állítom, hogy minden ízében friss még a szöveg és hogy helyenként nem haladta meg a tudomány Dobzhansky korabeli álláspontját. Ugyanakkor összességében még ma is igaz, amit leír, és fontos, amire reflektál. Hiszen egy olyan korban élünk, amikor megint nagyon is realisztikus, sőt kívánatos lett sokak számára, hogy a leendő generációk genetikai anyagában belenyúlva, a mi mostani ízlésük szerint alakítsuk azokat.
Előző évek könyvajánlói: 2022 | 2021 | 2020 | 2019 | 2018 | 2017 | 2016 | 2015 | 2014 – I. és II. | 2013
Mozambik 20. századi története magában hordozza a hasonló egykori gyarmatok összes nyomasztó jegyét. A brit magántársaságok önkényuralma után rövid német, majd hosszabb portugál fennhatóság következett és az 1975-ös függetlenség nem valami szebb jövő, hanem egy évtizedes pusztító polgárháború kezdete lett.
Utóbbi pedig nem kímélt senkit és semmit: nemcsak az emberéletek váltak beáldozható nüanszokká, hanem az állati életek is, különösen akkor, ha a puszta húsutánpótláson kívül esetleg bármi egyéb anyagi haszon is származhatott az adott élőlény lelövéséből. Talán semmi sem példázza azt jobban, mint a Gorongosa Nemzeti Parkban élő elefántpopuláció története.
A Nemzeti Park maga a Nagy-hasadékvölgy déli végénél elhelyezkedő terület, ami már a 1960-as évek óta nemzeti parkként működött és páratlan fajgazdagsága miatt a mozambiki függetlenség hajnalára méltán világhírű lett. A polgárháború aztán ezt a páratlan faj- és egyedgazdagságot teljesen eltüntette: az állományuk század, vagy ezredrészükre zsugorodtak és egyes fajok (pl. zebra és oroszlán) gyakorlatilag kipusztultak ezen a területen. Az elefánt populáció ehhez képest szinte szerencsésnek mondható, hiszen “csak” 90%-os csökkenést könyvelhetett el és az 1972-es 2542 egyed helyett 2000-re 242 állatot lehetett még fellelni. S ami még érdekesebb, ezzel párhuzamosan a populációban látványosan megnőtt az agyar nélküli egyedek aránya.
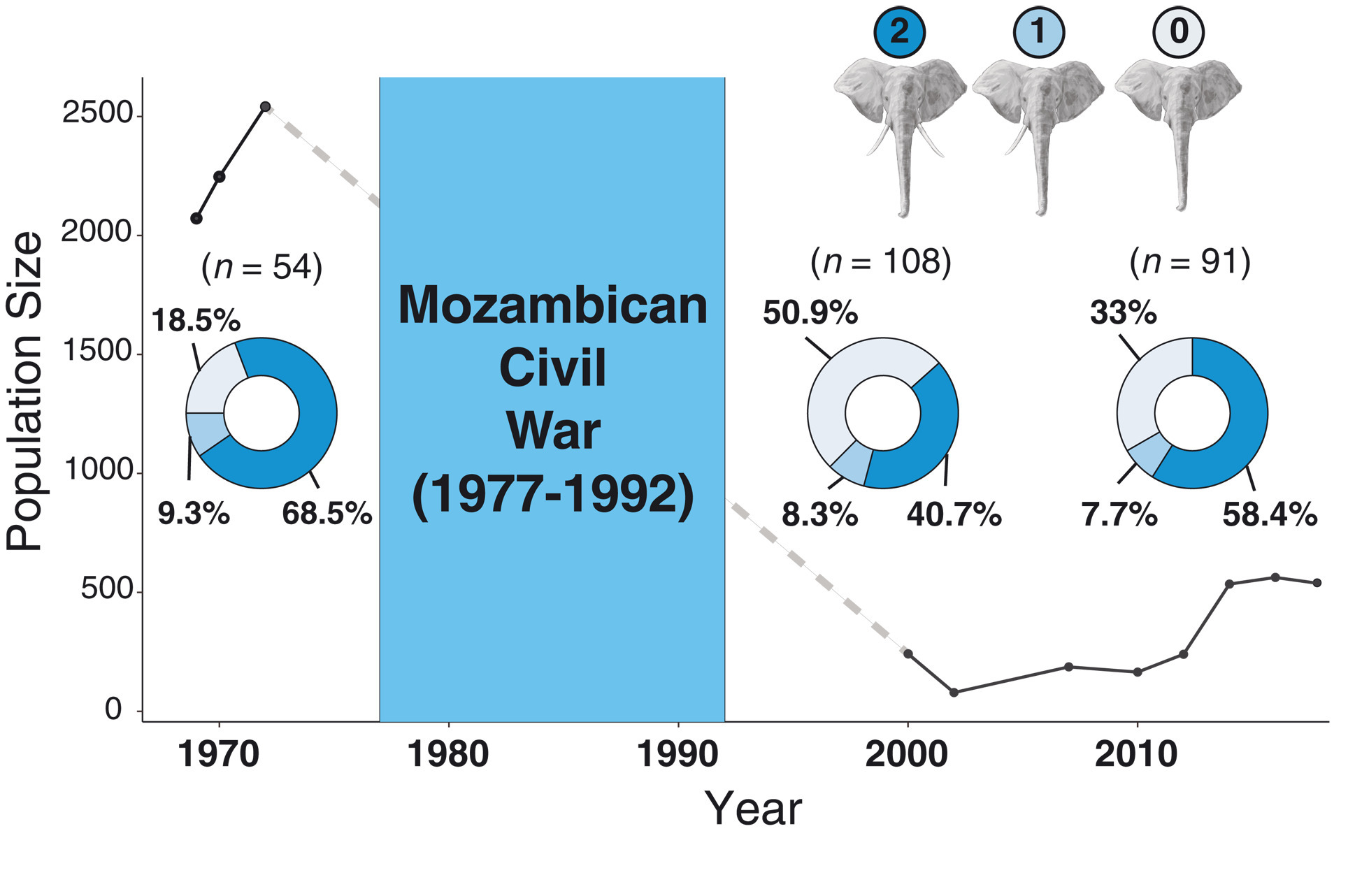
A Science-ben a tegnap megjelent cikk alapján az emberek által végrehajtott brutális szelekció eredménye egy olyan allél feldúsulása lehetett a populációban, ami nagyon szép példája a nemhez-kötött, letális domináns jellegek öröklődésének.
Először is nagyon érdekes, hogy a most már félévszázados elefánt-cenzusban, sosem lehet agyar nélküli hímeket fellelni, csak nőstényeket. Ez már önmagában is azt sugallja, hogy elképzelhető, hogy egy X kromoszómához kötött tulajdonságról van szó, amelynek agyartalnaságot okozó alléja (nevezzük ezt XD-nek) hímek esetében letális. Ha így van akkor az azt is jelentené, hogy az agyartalan nőstények pedig X+XD genotípusúak. Ez pedig két fontos predikcióhoz vezetne:

A megfigyelések ezt szinte tökéletesen hozzák, ezáltal egy új, szép de szomorú példát szolgáltatva a tankönyvek számára.
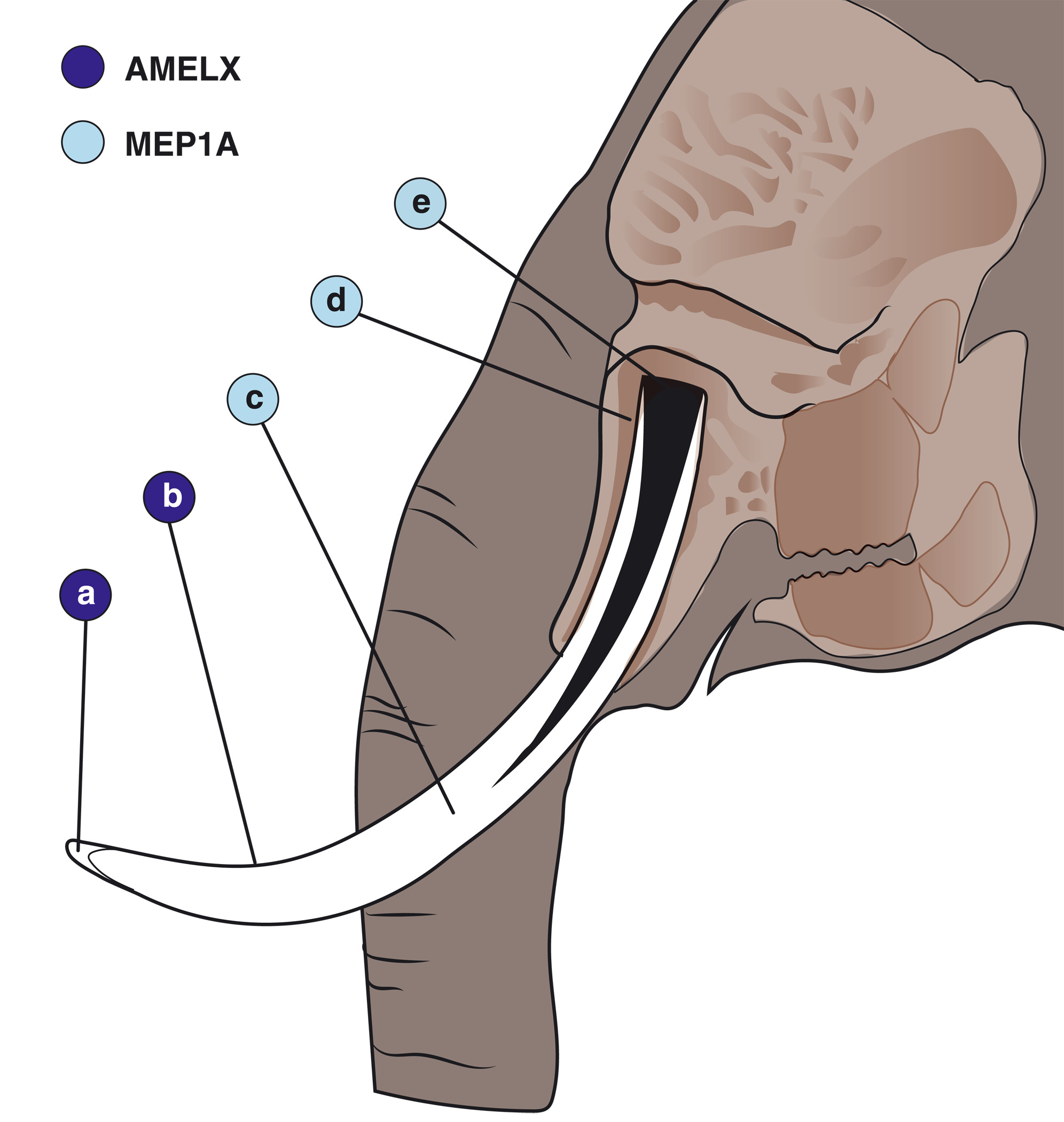
Persze fontos kérdés az is, hogy vajon mi is az a bizonyos génvariáns, aminek a nőstények agyartalansága köszönhető? Az eddigi vizsgálatok arra voltak elegendőek, hogy az X kromoszóma egy bizonyos szakaszára le lehessen szűkíteni a keresést. Itt egy AMELX nevű, a fog-fejlődésben szerepet játszó gén található. Utóbbi a zománcfejlődésben szerepet játszó amelogenin nevű fehérjét kódolja és emberekben az ezt tartalmazó régió deléciója egy zománcképződési problémákkal járó Amelogenesis Imperfecta nevű ritka betegséget okoz, valamint – nem mellesleg – hím letalitást.
A genom többi részét is megnézve, még látható szelekció nyoma az autoszomális (vagyis nem nemi kromoszómán levő) MEP1A gén környékén is, amely egy dentin mineralizációban lényeges faktort hoz létre.
Mivel a konkrét mutációt még nem találták meg, bizonyosságról nem beszélhetünk, de jelenleg az tűnik a legvalószínűnek, hogy az elefánt-agyar iránti kereslet egy olyan mesterséges szelekciós mechanizmust hozott létre, ahol az agyartalan elefántok szelekciós előnyhöz jutottak és így a populációban addig csak kis mértékben jelen levő mutáció egy ideig dominánssá tudott válni. A polgárháború vége és a nemzeti park helyzetének javulása lehetővé tette, hogy a természetes szelekció „vegye ismét kézbe” az allél-gyakoriságok szabályozását és talán pont ennek is köszönhető, hogy ismét az agyarakkal rendelkező állatok kerültek többségbe. (Nem zárható ui. ki, hogy az XD mutációnak egyéb olyan hatásai is vannak, amelyek a rátermettséget csökkentik – a cikkből sajnos nem derül ki például, hogy az agyartalan nőstények fogai is jobban megviselődnek-e.)
A Gorongosa Nemzeti Park egyébként azért is érdekes, mert az egész története reménykeltő, hogy egy kis odafigyeléssel teljes ökoszisztémákat vissza lehet hozni a megsemmisülés határáról. Erről szól (részben) egyik nagy kedvencem, Sean B. Carroll “Serengeti Rules” c. könyve, illetve az alább linkelt előadás.
(A borítókép forrása: howstuffworks.)
Campbell-Staton SC, Arnold BJ, Gonçalves D, Granli P, Poole J, Long RA, Pringle RM (2021) Ivory poaching and the rapid evolution of tusklessness in African elephants. Science 374(6566): 483-487. doi: 10.1126/science.abe7389.
Még a blog hajnalán futó, “Csökevényszervek” sorozatban írtam egyszer az emberi farokról, ami egyrészt azért érdekes, mert jobbára nincs, pedig az emberi fajt is magába fogadó rendszertani törzs, a Gerinchúrosok egyik legfontosabb bélyege az ún. posztanális farok, másérszt – ahogy az ominózus poszt is pedzegeti – néha, atavisztikus jegyként mégis feltűnik.
A teljes képhez még hozzá kell tennünk, hogy mindez igaz más emberszabásúakra is, illetve a farok tulajdonképpen elkezd kifejlődni embrionálisan, de aztán, jobbára a sejtek pusztulása (apoptózisa) miatt, az utolsó trimeszterben elsorvad.
Mindig is érdekes kérdésnek tűnt, hogy vajon mi okozhatta evolúciósan egy ilyen kulcsbélyeg eltűnését, miért volt erre szükség, illetve hogyan figyelhető meg mégis pár emberben csökevényes farok a születés után. Ezekből az első és utolsó kérdésre ad, ha nem is teljes, de részleges választ egy napokban a bioRxiv-on publikált preprint.
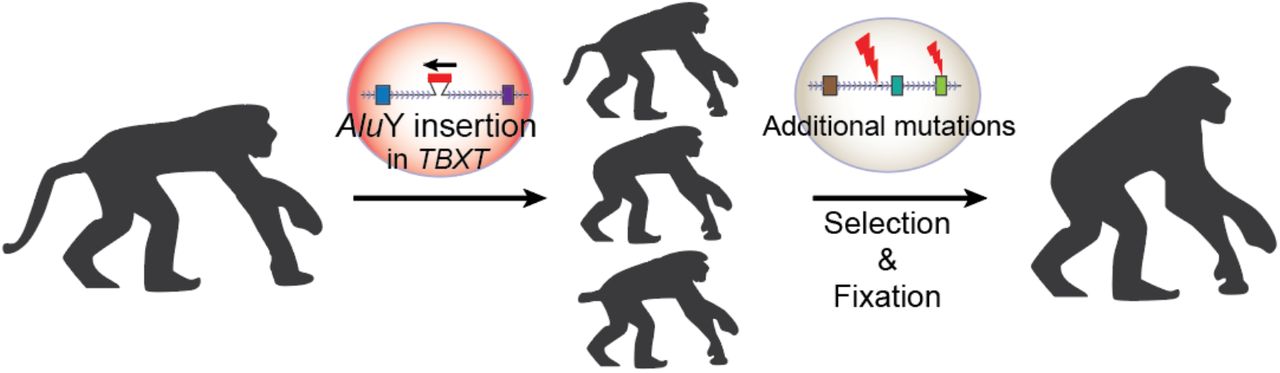
A szerzők fontos meglátása volt, hogy az emberi gerinc és farok fejlődésének egyik kulcsgénje, a TBXT esetében az emberszabásúak genomja egy fontos pontban eltér más főemlősökétől: a hatodik és hetedik exonok közt feltűnik egy új ugrálógén-kópia, az AluY.
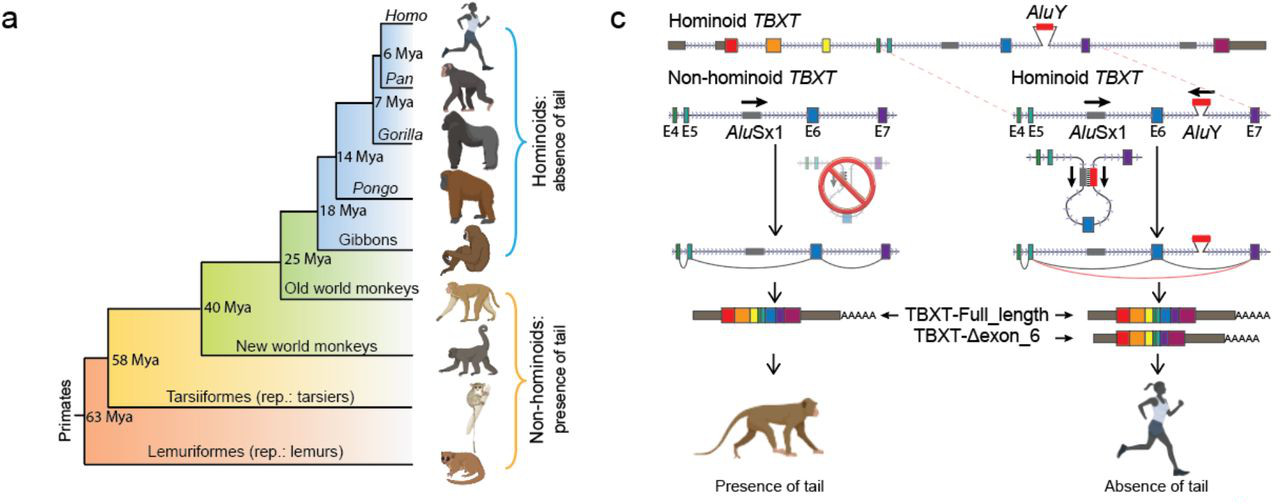
Az Alu elemek alkotják az emberi (pontosabban a főemlős) genom egyik legnagyobb “ugrálógén” közösségét, hiszen olyan, egyenként kb. 300 bp hosszú transzpozonokról van szó, amelyekből közel 1 millió van az emberi genomban, vagyis a genomunk több mint tizede ilyen szekvenciákból áll. Nem egy monolitikus transzpozontengerről van szó, ezeken belül is megkülönböztetünk különböző alcsaládokat, amelyek közül pár, így például a talán legfiatalabbnak tartott AluY is, még mindig aktívnak tűnik.
És ez adja a konkrét megfigyelés lényegét is, ugyanis a főemlősök genomjában a TBXT gén hatodik intronjában egy (evolúciósan) új AluY található meg. Ez azért vált különösen érdekessé, mert egy kicsit távolabbi rokona egy AluSx1 transzpozon az előző, vagyis ötödik intronban is felfedezhető és a két elem szekvenciahasonlósága kellően nagy, hogy a TBXT gén átíródásakor a még éretlen pre-mRNS-ben egy hurokszerű struktúra jöjjön létre, ami bezavar a pre-mRNS érésébe, azaz a splicing folyamatába.
Így emberben (és feltehetőleg más emberszabásúakban is) tulajdonképpen a TBXT gén két izoformája jelenik meg: a teljes hosszúságú és egy másik, amiből a hatodik exon hiányzik és egy csonkolt fehérje képződését segíti elő. Vagyis egy emberszabású embrió kis túlzással úgy viselkedik, mint egy heterozigóta mutáns (ahol szintén két verziója lenne jelen a génnek), s innen jött az ötlet, hogy mi lenne, ha meg is néznénk pont ezt egy olyan állatban, ahol egyébként kifejlődne a farok, pl. egy egérben.
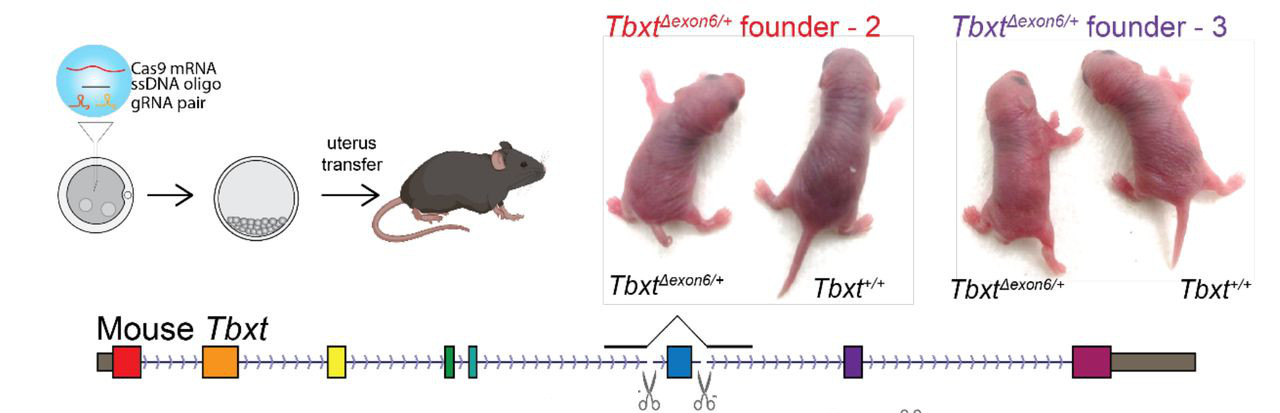
Modern genomszerkesztési technológiákat felhasználva létre is hoztak ilyen egereket és bingó, azoknak többnyire ilyen-olyan farok-fejlődési rendellenessége volt, többeknek pedig egyáltalán nem fejlődött ki a farka (összesen a hetrozigóták harmadánál volt valamilyen fenotípus). Ezeket a kísérleteket még kiegészítették emberi sejtvonalakon végzettek is, amelyek szintén visszaigazolták, amit sejteni lehetett: mind az AluY, mind az AluSx1 szükséges a plusz izoforma kialakulásához, hiszen egyik, vagy másik hiányában nem alakul ki a pre-mRNS érésbe bezavaró hurok-struktúra.
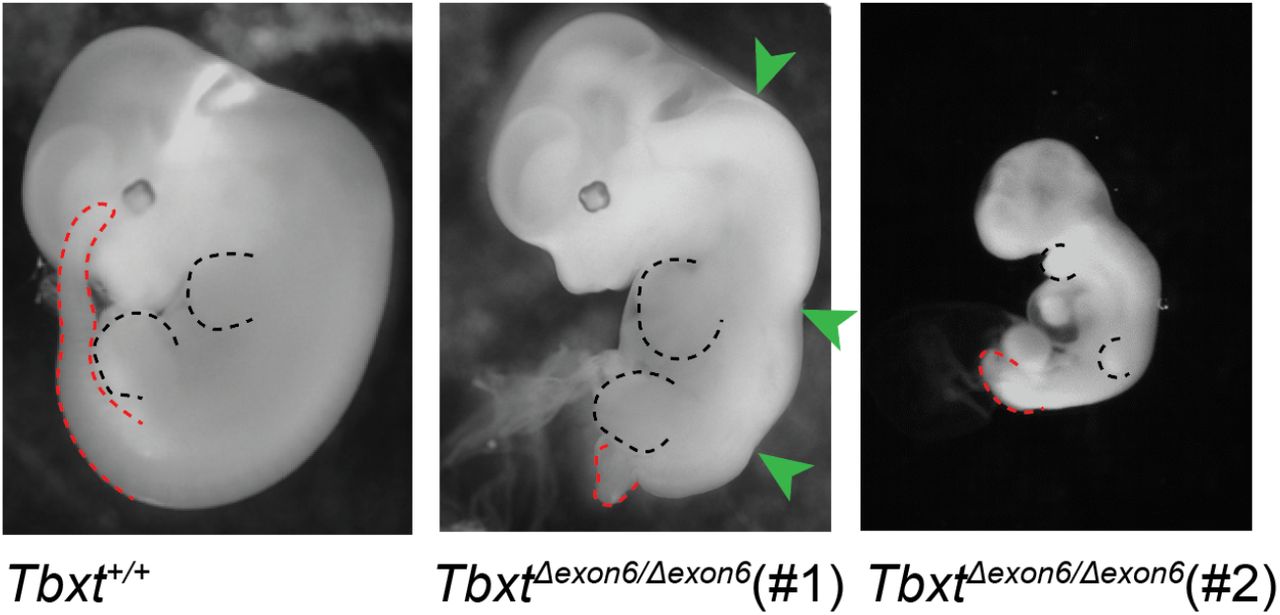
A mutáns egereknek köszönhetően azt is tudjuk, hogy a hatodik exon deléciója homozigóta állapotban halálos és az ilyen egerek a farok hiánya mellett az emberi nyitott gerincre (spina bifida) emlékeztető elváltozásokat mutattak.
Összességében tehát úgy tűnik, hogy az emberszabásúak faroktalanságát egy transzpozon túlbuzgóságának köszönhetjük, amelynek következtében a TBXT gén két izoformája is jelen van a sejtekben. Valamiért (s itt azért még tapogatózunk) a faroknélküliség egy előnyt jelenthetett a földön több időt töltő őseinknek és megjelenhettek és rögzülhettek olyan stabilizáló mutációk is, amelyek a csak részben penetráns faroktalan fenotípust (a heterozigóta egerek csak harmada lesz az), teljesen penetránssá tették. Ugyanakkor, ha valamilyen egyéb okokból változik a két TBXT izoforma aránya, jó eséllyel annak vagy az lesz a következménye, hogy nyitott gerincű gyerekek születnek, vagy épp olyanok, akiken ektopikus, atavisztikus farkak jelennek meg.
(A fedőkép a Wikimedia oldaláról származik.)
Xia B, Zhang W, Wudzinska A, Huang E, Brosh R et al. (2021) The genetic basis of tail-loss evolution in humans and apes. bioRxiv 2021.09.14.460388; doi: 10.1101/2021.09.14.460388
A napokban jelent meg egy régóta dédelgetett, Kun Ádám-mal közösen írt cikkünk a Scientific Reports-ban. Gondoltam írok róla egy „akadémiai hokedlifoglalót”, hátha elő tudom adni azt érdekesen. Még akár akkor is, ha az olvasó nem biológus. Szemtelen önmarketing következik…
Akad pár hasonlóság az élő szervezetek és civilizációnk vívmányai között. Én napjaimat azzal töltöm, hogy betűk formájában vetem papírra a fejemben keringő gondolatokat. Ehhez a billentyűzet megfelelő gombjait kell lenyomnom egymás után. Ezekből a betűkből végül szavak, és mondatok lesznek.
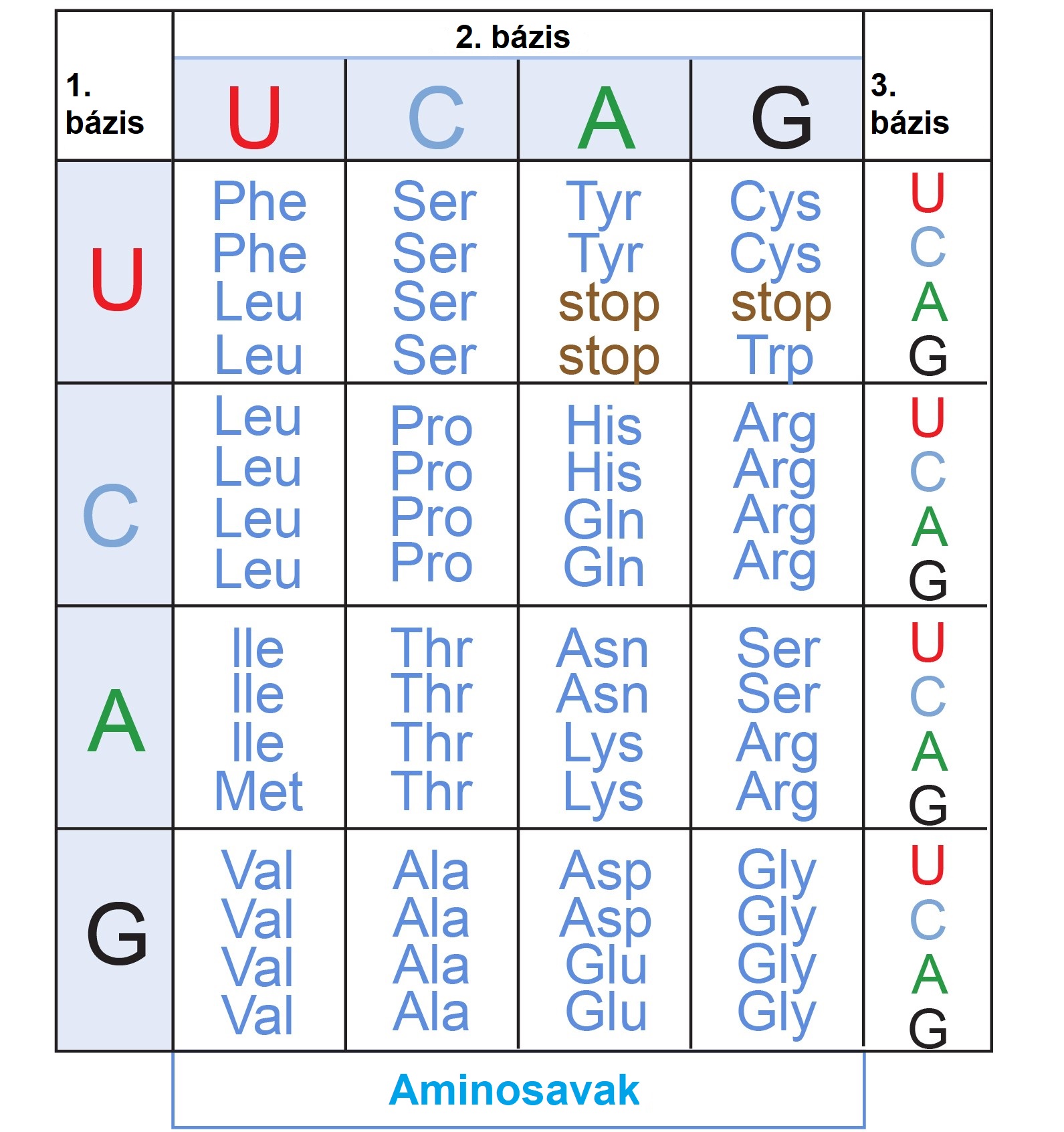
Ugyanilyen beleéléssel püföli saját kis genetikai billentyűzetét a Föld összes organizmusa, hogy a DNS-ben található, kódolt információt lefordítsa az életfolyamatokhoz nélkülözhetetlen fehérjékre. Ebben az esetben a DNS-en hordozott bázishármasok (pl. AGC, CAA) határoznak meg egy aminosav-sorrendet, mely a fehérjéket alkotja.
A hasonlóság itt nem ér véget. Az emberek túlnyomó többsége ugyanis majdnem ugyanazt a „QWERTY” billentyűkiosztást, vagy annak variánsát használja (1. ábra; a magyar QWERTZ billentyűzet ezzel szinte teljes mértékben megegyezik). Ennek egyik következménye, hogy ékezetektől eltekintve vakon képesek lennénk ugyanazt a szöveget bepötyögni bárhol. Az adott karakterek pozíciója nem változik a klaviatúrán, legyen szó számítógépről vagy telefononról, Svédországtól egészen Kínáig.

Ehhez hasonlóan minden organizmusban ugyanaz a genetikai ”billentyűkiosztás” található meg! Ha fogok egy emberi DNS-darabot, és azt egy másik élőlénybe, például egy baktériumba ültetem át, megkaphatjuk ugyanazt az fehérjét, ami bennem is termelődik. Egy adott bázishármas, csakúgy mint egy leütés helye a klaviatúrán, ugyanarra a karakterre (aminosavra), fog dekódolódni, pl. CCA → prolin, AGG → arginin (2. ábra). Mindenkiben ugyanaz a fordítócsomag, ugyanaz a genetikai kód fut.
Hiába hasonlít a két példa csak felületesen egymásra, mindkettő felvet egy közös kérdést:
Miért használ mindenki QWERTY billentyűzetet és miért pont ez standard genetikai kód található meg a Föld valamennyi élőlényében? Milliárdnyi más felosztást el tudunk képzelni. Például a billentyűzeten valaki igazán sorba is rendezhette volna a betűket, mégse azt használjuk. Hasonlóan máig nem tudjuk, hogy miért pont ez egy genetikai billentyűzet terjedt el a Földön és ez miként alakult ki több milliárd évvel ezelőtt. Ez a Földi élet keletkezésének egy kulcskérdése. A kirakós egyik darabkájának feltárásában lesz segítségünkre ha megfejtjük az emberi billentyűzetet.
Magát a QWERTY-kiosztást még az 1870-es években fejlesztette ki Christopher Latham Sholes. Tervezése a kényelmes, lehető leghatékonyabb gépeléshez lett igazítva. Ehhez a legyakoribb karaktereknek és karakterkombinációknak könnyen kézre kell esniük, s ez az ist jelenti hogy gépelés során az egymást követő betűk leütéséhez a lehető legkisebb távolságot kell megtennie az ujjaknak, mindezt a lehető leggyorsabban.
Ugyanakkor a karakterek gyakorisága függ a beszélt nyelvtől is! Jelentős eltéréseket találhatunk az egyes nyelvek között (3. ábra). Például a kínai pinjin (a mandarin latin betűs átirata) nagyon elüt az európai nyelvektől ebben a tekintetben. Nem mindegy tehát hogy milyen nyelven gépelünk a QWERTY billentyűzeten.

Ezek a különbségek kihathatnak a gépelés hatékonyságára is. Ezt akár mi is könnyűszerrel ellenőrizhetjük (gép előtt ülők előnyben). Az angolban a leggyakoribb egymást követő karakterpárok: t-h, h-g, i-o, míg a pinjinben: z-h, c-h, s-h. Csak nézzük meg mekkorák a távolságok az egyes billentyűk között!
De ha ez nem lenne elég, akár tovább is mehetünk. Kiszámolható, és kísérletesen ellenőrizhető ugyanis, hogy milyen hatékony a QWERTY billentyűzet az egyes nyelvek esetén (4. ábra). Jól látható, hogy az billentyűk közti távolság az angolban és a hozzá valamelyest hasonlító németben a legkisebb. Őket követi a francia és a spanyol. A kínai pinjin-ben sokkal nagyobb távolságokat kell megtenni leütések közben, ez pedig azt is eredményezi hogy ezen a nyelven sokkal lassabban lehet csak gépelni.
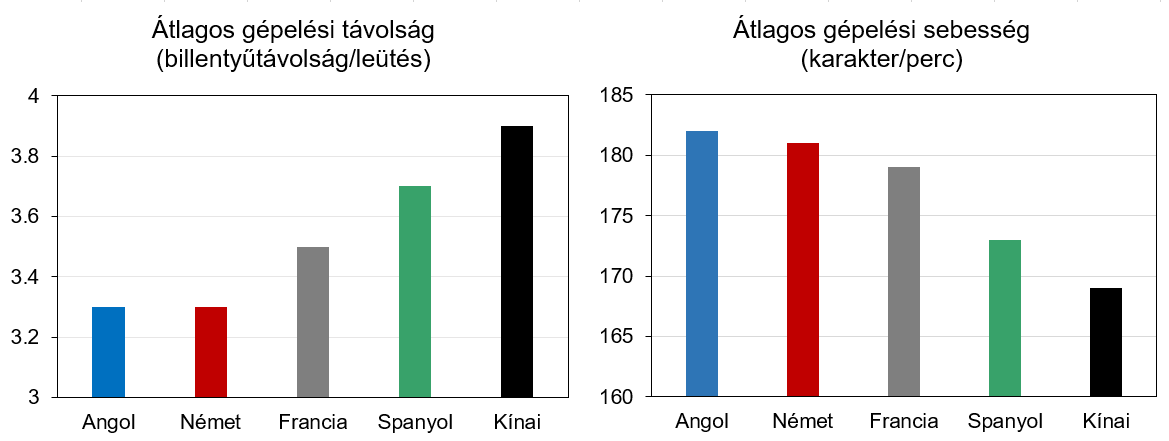
Az okot valószínűleg már az olvasó is gyanítja: e különbségek elsődleges oka az, hogy a QWERTY-kiosztást amerikaiak tervezték, az angol nyelv igényeire szabva. Minél kevésbé hasonlít egy nyelv az angolra, az annál inkább küszködni fog ezzel az univerzális billentyűzettel. A mi kutatásunk egy hasonló gondolatmenetet követett a genetikai kód eredetének feltárásához.
Cikkünk egy hasonló jelenség megfigyelése volt a genetikai kóddal kapcsolatban. Azt már sejtjük egy ideje, hogy a genetikai kód valamelyest optimalizálódott a fehérjék gyártásához. Ennek vannak egyértelmű jelei. Például régóta ismert, hogy a genetikai kód próbálja csökkenteni a „félreütés” kockázatát: ha történik egy mutáció a DNS-ben, az továbbra is ugyanazt, vagy hasonló tulajdonságú aminosavat fog eredményezni a fehérjében. Ezzel pedig lesz rá esély, hogy a „félregépelt” fehérje továbbra is működőképes marad.
Az egységes kód ellenére azonban az élőlények is különböző „nyelvjárásokat” használhatnak, más gyakorisággal püfölik e genetikai billentyűzet gombjait (5. ábra). Az eltérés részben a környezet függvénye. Egy bélbaktérium (Escherichia coli) más összetételű fehérjéket igényel, mint pokoli körülményeket is tűrő mikrobák, akik akár még forráspont körüli termálvizekben is vidáman lubickolnak (pl.: Thermus thermophilus és Ignisphaera aggregans).
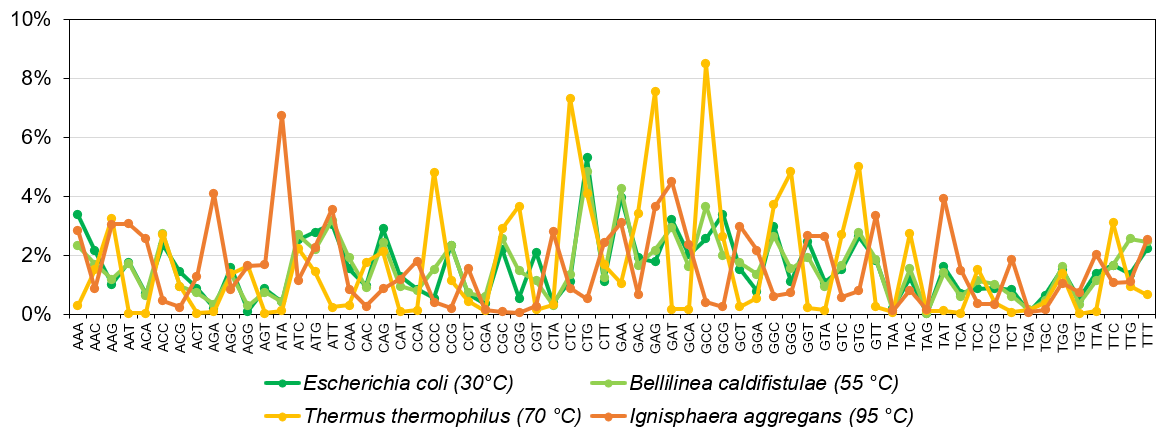
Noha a különbségek nem nagyobbak mint irodalmi magyar és a szögedi ömbörök böszéde közti eltérés, az mégis kihathat genetikai kód hatékonyságára. Mi többek között azt mutattuk ki, hogy a forróbb élőhelyeket meghódító organizmusok e téren igenis alulmaradnak a hűvösebb környezetekben éldegélő „szobabacikkal” szemben (6. ábra). A standard genetikai kódot használva nagyobb a mutációk költsége az ún. extremofil organizmusokban, s így ők nem tudnak olyan hatékonyan gépelni.
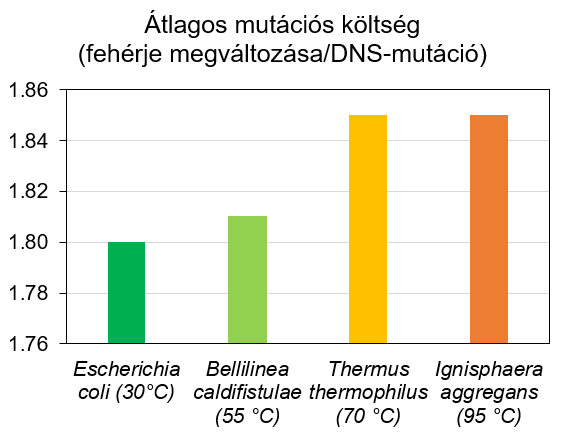
Mindez akár azt is jelenheti, hogy a genetikai kód jelenlegi formája nem biztos hogy extrém forró körülményekre lett kifejlesztve. Sokkal inkább egy enyhe, tavaszias időjárás képe bontakozik ki. Ez részben meghökkentő. Előszeretettel szeretjük azt gondolni, hogy hajdanán az Élet valami Yellowstone Nemzeti Park-szerű világban látott napvilágot, netán mélyvízi vulkanikus füstölgőkből kúszott elő. Én személy szerint mindig úgy képzeltem el az Élet bölcsőjét mint Csodaturmix varázsfőzetét forrni az üstben.
Azonban korántsem lehetetlen az sem, hogy valamikor hűvösebb körülmények uralkodtak a Földön, és ez adott otthont az első mikroorganizmusoknak. Az viszont, hogy ennek forgatókönyvnek a jelentőségét tisztázzuk és komolyan alátámaszthassunk, már a jövő zenéje…
És természetesen ez nem jelenti azt, hogy sikerült minden kérdésre választ találni a genetikai kóddal kapcsolatban. De lett egy új puzzle darabkánk, aminek segítségével több esélyünk lesz kirakni egy teljesebb képet az Élet keletkezésének korai fázisairól.
Források:
Radványi Á. & Kun Á. (2021) Phylogenetic analysis of mutational robustness based on codon usage supports that the standard genetic code does not prefer extreme environments. Sci Rep 11, 10963.
Bi X., Smith B. A. & Zhai S. (2012) Multilingual touchscreen keyboard design and optimization. Human–Computer Interact 27, 352–382.
 Ha Darwinról és hozzá közel álló madarakról beszélünk, akkor talán nem is feltétlenül a kötelező újggyakorlatként felemlegetett galapagosi pintyeket (aka. “Darwin pintyek”) kellene előhozzuk, hanem a galambokat.
Ha Darwinról és hozzá közel álló madarakról beszélünk, akkor talán nem is feltétlenül a kötelező újggyakorlatként felemlegetett galapagosi pintyeket (aka. “Darwin pintyek”) kellene előhozzuk, hanem a galambokat.
Nem sokkal “A fajok eredete” második kiadását követően (amiből szintén nem hiányoztak a galambok) publikálta Darwin „Az állatok és növények megváltozása a háziasítás állapotában” (The Variation of Animals and Plants under Domestication) c. könyvét, amiben két teljes fejezetet szentelt a különböző galambfajtáknak. Figyelembe véve, hogy Darwin maga is lelkes űzője volt kora állattenyésztési hobbijának, mindez talán nem is annyira meglepő, talán inkább az a releváns kérdés, hogy mennyire formálta a galambtenyésztés Darwin nézeteit az evolúcióról és hogyan használta kvázi modellélőlényként galambjait.
Mindenesetre, ha nem is pont így fogalmazott, Darwint saját keresztezései győzték meg arról, hogy a kedveltebb fajták kinézete tulajdonképpen olyan ritka természetes variációknak köszönhető, amelyek egyébként is jelen lehettek az ősi galambokban. És bár Darwin értelemszerűen még nem használta a domináns és recesszív fogalmakat, az általa leírt egyik kísérlet, amiben három fajtiszta vonal különböző hibridjeit keresztezte és végül az ősi formának tekintett szirti galambra emlékeztető utódok jöttek létre, tökéletesen példázza, hogy mi is történne, ha különböző gének recesszív alléljaira homozigóta (és a normálistól eltérő kinézetű) vonalakat egymással kereszteznénk.
Bár a galambászati kedv napjainkra kissé alább hagyott, azért még mindig kellő számban léteznek ezek a több száz év alatt létrehozott formák, hogy tanulmányozni tudjuk, mi is hozza létre a jellegzetes fenotípusos jegyeket, például a parókás galamb csuklyáját, vagy az angol trombitás tollas lábait. (Egyébként a mesterséges szelekció létrehozta formagazdagság, csak a kutyában és az aranyhalban tapasztalthoz hasonlítható, vagyis két olyan másik fajhoz, amelyeket szintén évszázadokon keresztül formált saját ízlése szerint az ember.)

Utóbbi azért is érdekes téma, mert a blog történetének egyik leggyakrabban elővett témájához vezet (ismét) vissza minket, ez pedig a végtagfejlődés. De menjünk sorban.
Az elmúlt években a galambok evolúciós modellélőlényként való felfedezése elsősorban a University of Utah-on dolgozó Michael Shapiro nevéhez köthető. Az ő csoportja volt például az, amelyik öt évvel ezelőtt egy borzasztó érdekes eLife cikkben a galambok (és tyúkok) tollas lábainak fejlődési eredetét próbálta feltárni.
Egy átlagos madárláb, ahogy azt mind tudjuk, pikkelyes, karmos – és szöges ellentétben az állatok mellső végtagjával, jellegzetesen tolltalan. De akkor mi történhetett mondjuk az angol trombitás esetében? Hogy megtaláljuk melyik gének lehetnek a fenotípus mögött, Shapiroék az egyik klasszikus, már Mendel által is alkalmazott “trükköt” vetették be: keresztezték a különböző kinézetű (fenotípusú) madarakat, létrehozhva egy hibrid generációt (F1), amelyeket egymás közt szaporítva immár egy olyan generációt kaphattak (F2), ahol a recesszív jelelgek is ismét meg tudtak nyilvánulni.
A modern genomika egyik előnye egyébként az is, hogy elég pontosan lehetett azt is látni, hogy melyek azok a DNS szakaszok, amelyek a két kiindulási fajta genomjának elvileg teljes összekeverése ellenére minden egyes tollaslábú F2 egyedben jelen lehetnek és megegyezenek a tollas lábakkal rendelkező kiindulási szülő esetében. Ezek rejtik jó eséllyel ugyanis azokat a mutációkat/szekvencia-variánsokat, amelyeknek köze lehet a furcsa lábak kialakulásához.
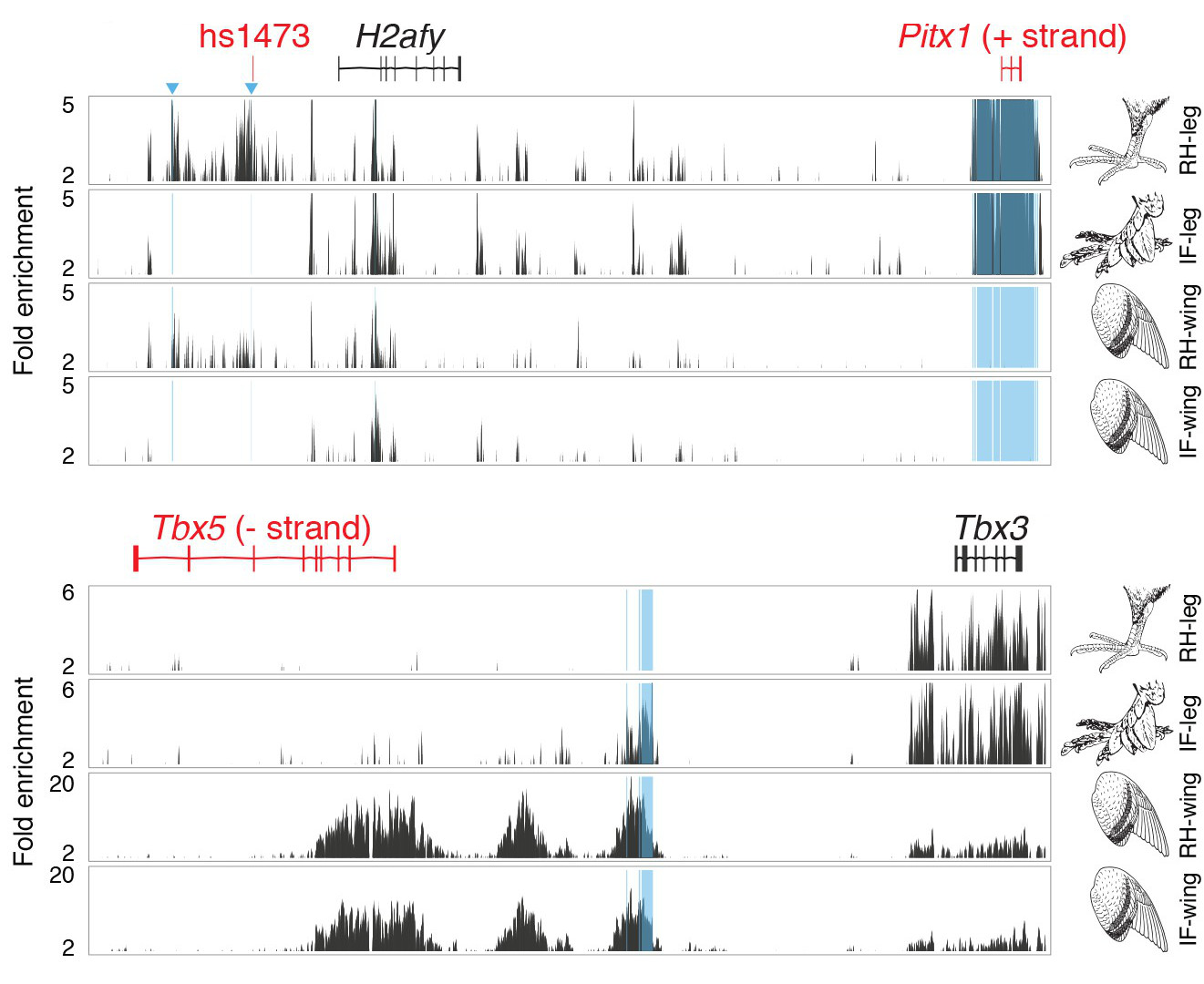
Ennek az eredményeként két fontos genomi pozíciót leltek, amelyek ráadásul a végtagfejlődésben eleve fontos gének közelében vannak: az egyik a Pitx1 gén mellett, a másik a Tbx5 és Tbx3 gének között helyezkedett el. Hogy ez miért felel meg kvázi a jackpot-megütésének egy ilyen típusú tanulmányban, ahhoz egyetlen sorát venném elő a már emlegetett végtagfejlődéses klasszikus posztnak:
A mezodermális szövet jellegét pedig három transzkripciós faktor határozza meg: a mellső végtagért a Tbx5, míg a hátulsóért a Tbx4 és a Pitx1 a felelős.
Azaz két olyan gén közelében láthatunk változásokat, amelyek terméke pont azt szabályozza, hogy a mellső és hátsó végtagok különbözőek legyenek. És a jelek szerint ez a mutációk hatására épp ez a különbség kezdett elmosódni a tollas lábak esetében.
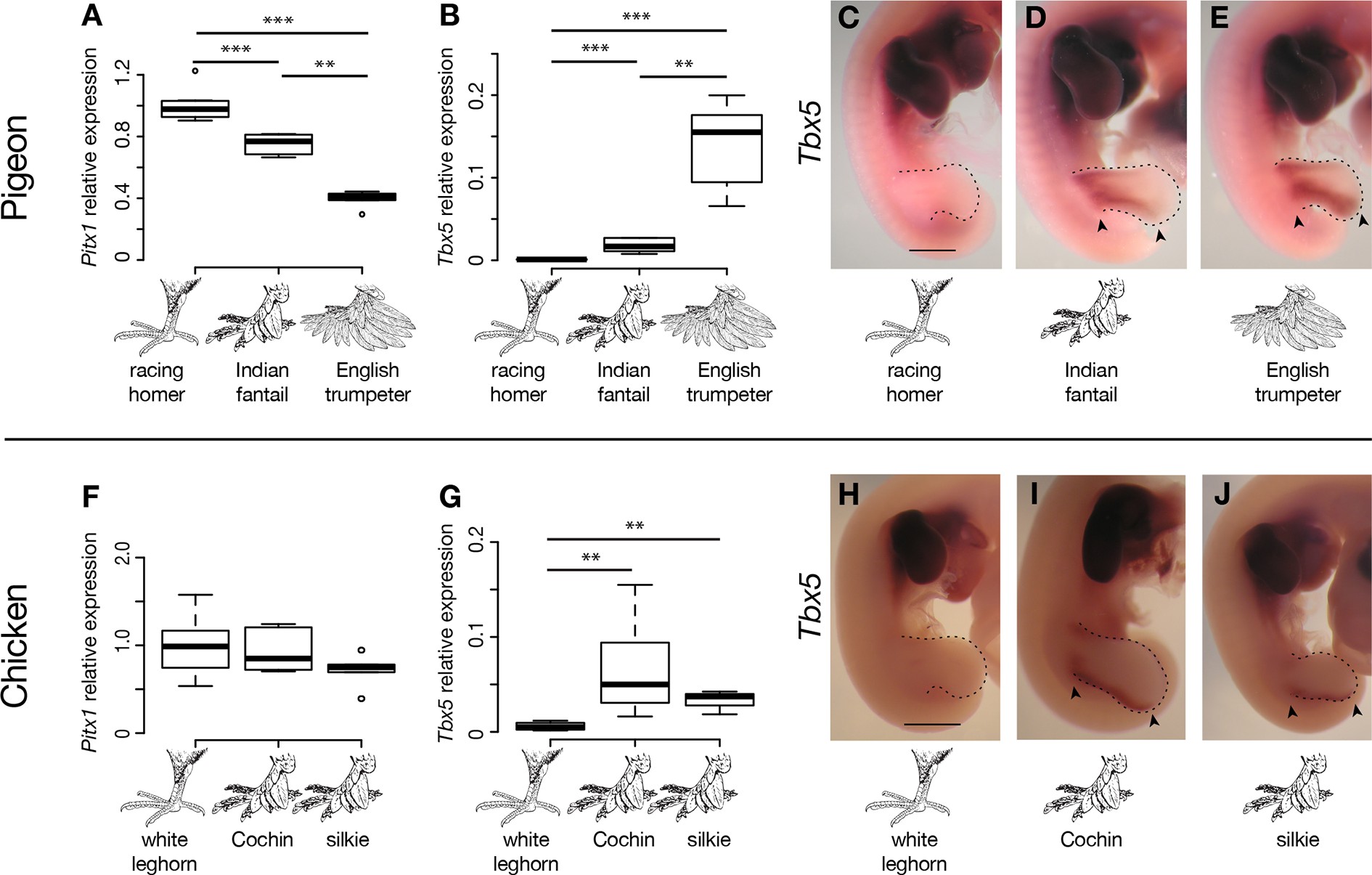
Ahogy a fenti ábra is jól mutatja: az angol trombitás esetében a hátsó végtag identitását biztosító Pitx1 expressziós szintje lecsökken, míg a normálisan csak a leendő szárnyakban expresszálódó Tbx5 hirtelen a hátsó végtagbimbóban is elkezd kifejeződni a fejlődés során. (S bár előbbi változás nem látható a gatyás tyúkokban, utóbbi annál is inkább.)
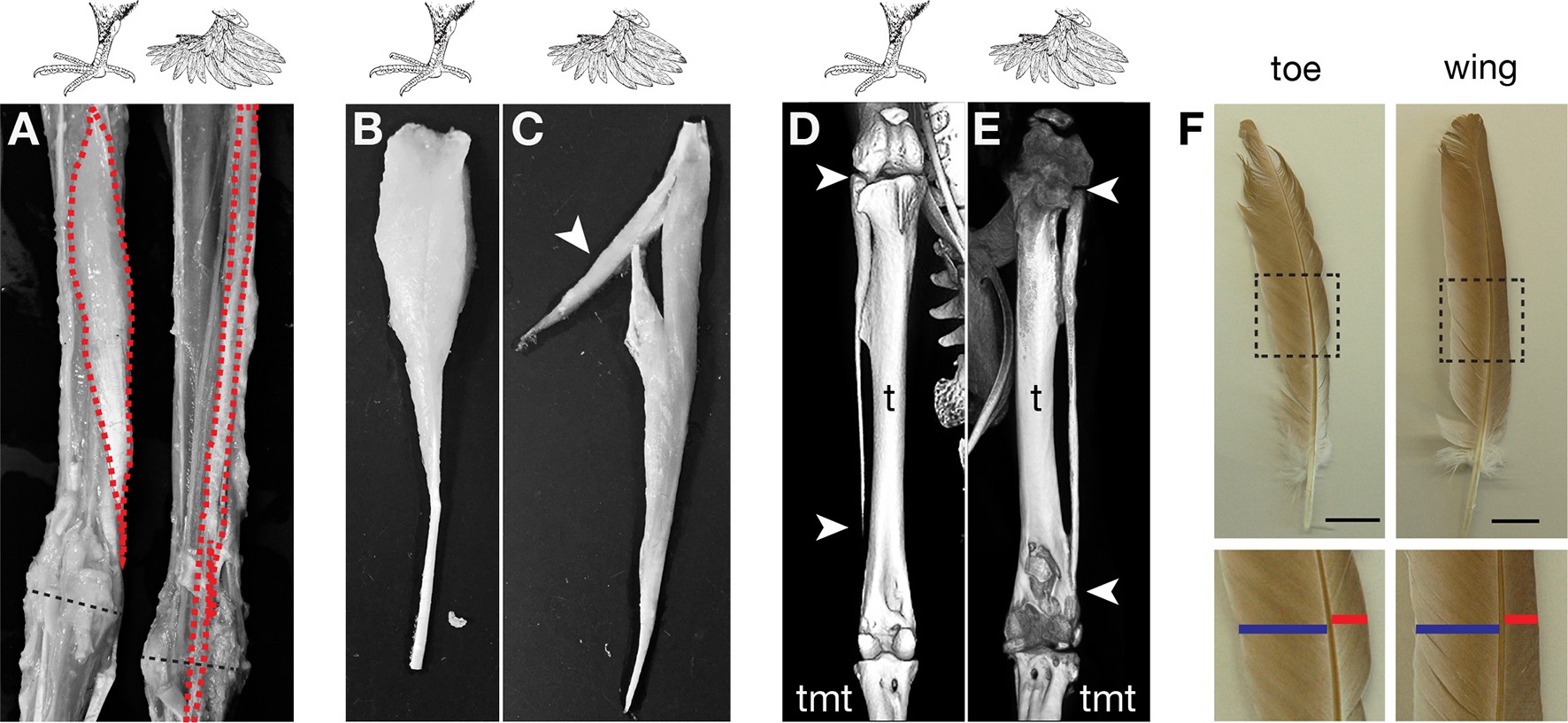
Ennek az ismeretnek a birtokában már talán kevésbé meglepő, hogy ha tüzetesebben megnézzük a tollas lábú galambok lábait, akkor egyrészt azt fedezhetjük fel, hogy ezek nem egyszerűen tollasok (és nem random tollakat, hanem a szárnyakra is jellemző fedőtollakat hordoznak), de alapvetően különböznek egy “normális” lábtól. Talán túlzás lenne azt állítani, hogy teljes egészében szárnyakká alakultak, de mindenképpen jól látható, hogy mind a csontok, mind az izmok alakja jellegzetesen különbözik a klasszikus madárlábtól, így egy kicsit “átmeneti” morfológiával rendelkező végtagot találunk itt és minden jel szerint ez az identitásvesztés, ami a tollazat megjelenését hozta magával.
Shapiroék csoportja azonban nem állt meg itt és pár további galamb-jelleg térképezésébe is belevágott, s ezekről kerültek fel preprintek a bioRxiv felületére.
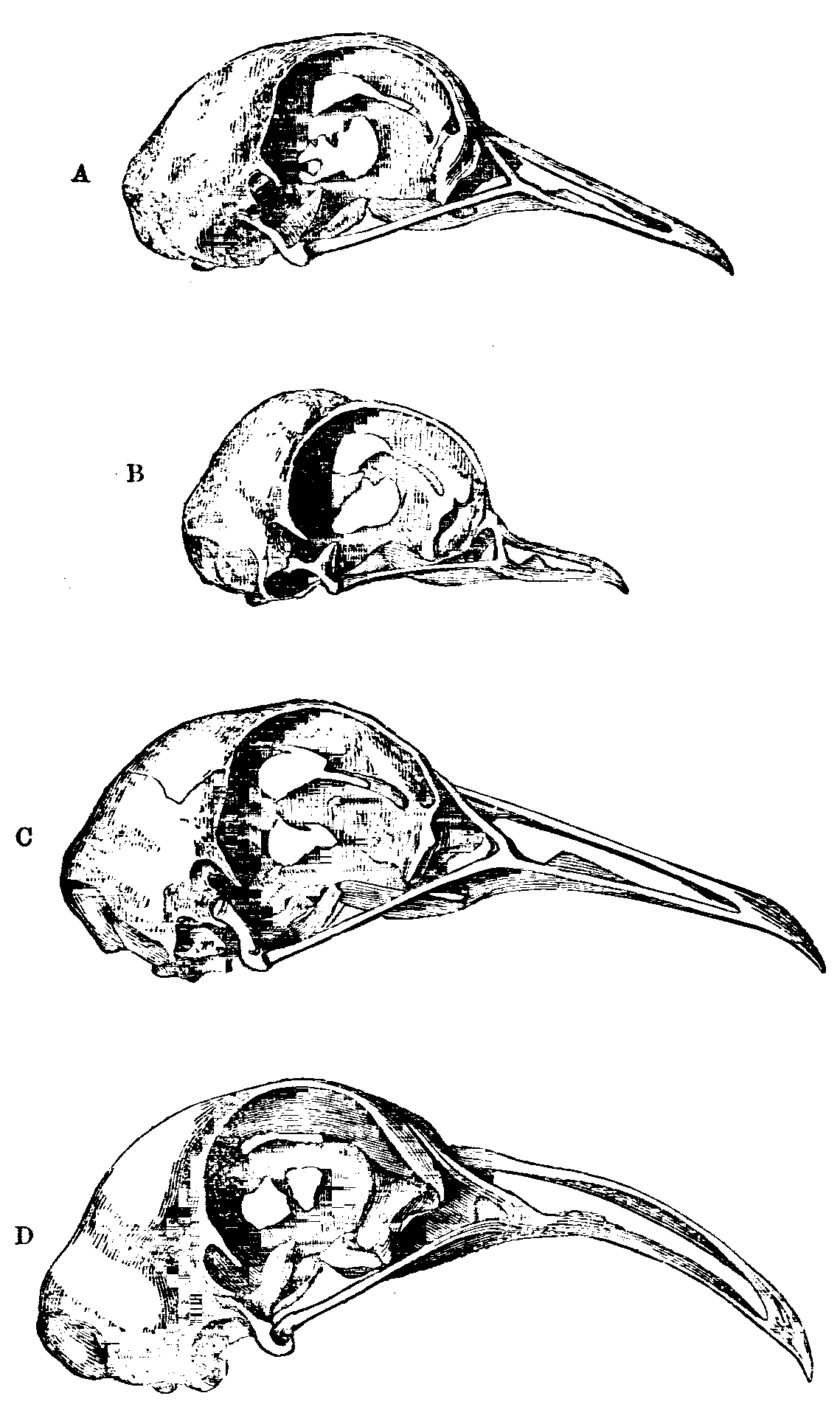
Az elsőben egy kicsit a galambkoponya alakjának furcsaságait próbálták megérteni. Ez egyébként szintén Darwin szívéhez közel álló téma, hiszen a „Az állatok és növények megváltozásában” maga is hosszan elemezgette ezt a dolgot.
A galambászat kiinduló fajának tekintett szirti galamb koponyájához képest, számod domesztikált fajtában, például az angol rövidcsőrű galamb esetében, látványosan megváltozott a koponya és csőr alakja és méretaránya. De talán sehol nem annyira látványos ez, mint az alsó állkapocs esetében, ahol akkora különbségek láthatók mondjuk a berber galambok, vagy az óriásra növő runtok esetében, hogy Darwin szavaival élve:
„Senki sem hitte volna el egykönnyen, hogy azok az állkapcsok, amelyek a fentebb meghatározott pontokban ennyire különböznek, egyazon fajhoz tartozhattak.”
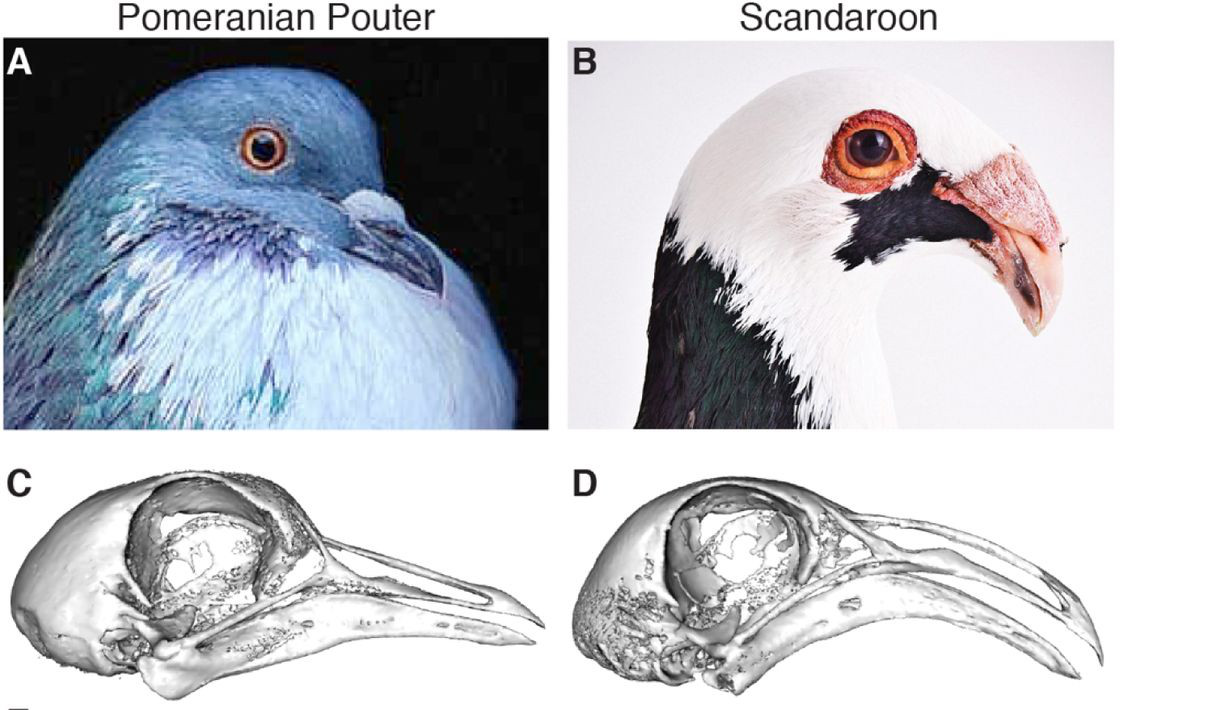
Vagyis Shapiro csoportjának bőven volt miből válogatnia. Végül egy pomerániai begyes galambot és egy scandaroont kereszteztek, mert ezek csőrmorfológiája kellően különbözött, hogy egy kellően részletes jellemzéssel az F2 generációban már nagyon sok csőr-paraméter genetikai hátterét fel lehessen tárni.
A minél pontosabb jellemzéshez Shapiróék csoportja mikro-CT készülékek segítségével beszkennelte a madarak koponyáját és nagyon egzakt módon rengeteg paraméterét lemérték. S bár egyelőre, a tollas lábakhoz kapcsolódó tanulmányokkal ellentétben, ezek eredményeként nem tudunk konkrét géneket megnevezni, azért viszonylag pontosan meg lehet mondani, hogy melyik kromoszóma-régiókban rejtőznek a minket érdeklő genetikai variációk. (Ennek a metodológiáját valahogy úgy kell elképzelni, hogy néhány tucat F2 hibridnek megszekvenálják a genomját és lemérik a koponyáját a CT segítségével. Ezt követően pedig elkezdenek korrelációkat keresni bizonyos markerek jelenléte és bizonyos fizikai paraméterek között. Mivel a koponya- és csőrforma egyértelműen genetikailag kódolt, ilyeneknek kell lenniük. A kérdés ilyenkor inkább az, hogy elegendő-e ennyi egyed megszekvenálása, hogy ezeket mind megleljük – általánosságban, minél több gén befolyásol egy karaktert, annál több egyed szekvenciájára és méreteire van szükségünk az ilyen vizsgálatokban, hogy “észrevegyük”, hol bujkálnak a releváns mutációk.)
A koponya szélességének és hosszúságának legfontosabb paramétereit öt kromoszóma-régió befolyásolja, a koponya formáját pedig további hat-nyolc. Ez elég sok (például a galapagosi pintyeknél úgy néz ki, hogy csak két–három gén játszik szerepet a csőr evolúciójában), különösen mert a mostani adatok nem képesek egyetlen génre leszűkíteni a keresést, hanem kromozsóma szakaszokat tudnak csak megadni. És jelen pillanatban egy-egy ilyen régió akár több tucat gént is tartalmazhat, amelyek közül a legtöbb (még) nem lett sehogy a koponyaforma evolúciójához kötve más fajokban. Így aztán, bár részletes felsorolás adható, a szerzők is csak találgatni tudnak, hogy melyik génnek van valójában köze a csőr és koponya változásaihoz.
Ennél szerencsésebb a helyzet a szivárványhártya (írisz) esetében, ami normális esetben narancssárga színű a szirti galambnál, de vannak fajták, ahol gyöngyházszínű és megint máshol pedig színtelen, ami miatt a mögötte található retina fekete pigmentált epitéliuma adja majd a (fekete) színt.
A furcsa színek megértéséhez tudnunk kell, hogy a szemben három fajta pigment van jelen: a fekete melanin (a retinában), illetve a fehéres guanidin és a narancssárga pteridin a szivárványhártyában. Minden pigmens több lépéses bioszintézis útvonalak állítanak elő, melyeknél elég, ha az egyik elem megsérül és az adott pigmens (illetve az általa meghatározott szín) is eltűnik.

A galamboknál különböző keresztezések alapján úgy tűnik, hogy mind gyöngyházszínű (pearl), mind a fekete (bull) szemszínt valamilyen viszonylag egyszerű, monogénes jellegként lehet vizsgálni.
Ennek alapján narancssárga és gyöngyházszínű szemű szülők unokáit (F2) vizsgálva kiderült, hogy az okozati mutáció jó eséllyel egy SLC2A11B nevű génben keresendő, ugyanis a fehéres szemű egyedekben ez a gén egy olyan mutációt hordoz, ami miatt terméke rövidebb és működésképtelen lesz. Viszonylag keveset tudunk erről a konkrét génről, eltekintve, hogy a sejtmembránban levő szállító funkciót betöltő fehérjét kódol és az ilyen típusú géneknek egyébként is szerepe van mindenféle állat pigmentációjában (igen, a miénkben is), a medaka nevű ázsiai halfajban konkrétan a pteridin-szintézishez is köze lehet.
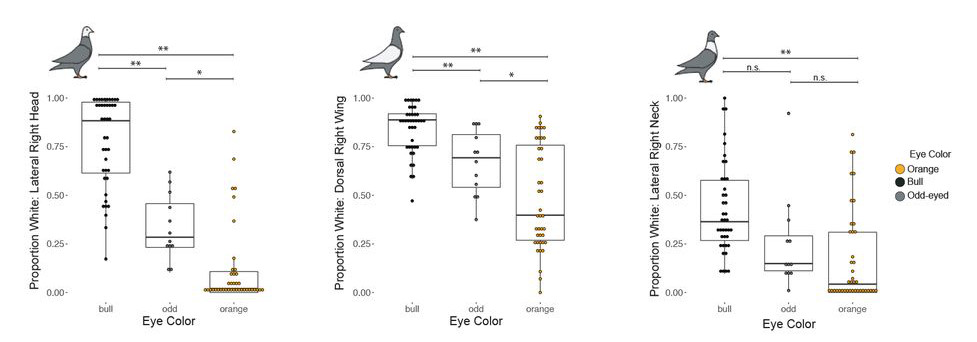
De a SLC2A11B csak fela a történetnek, a pigmentáció teljes hiányát a bull-szemű egyedekben más génnek a számlájára kell/lehet írni. Ez pedig, legavlószínűbb, hogy az EDNRB2 nevű, endothelin-receptort kódoló gén lehet. Tény, hogy itt nem azonosítottak olyan mutációt a kutatók a fekete szemű egyedekben, ami a gén működésképtelenségére utalna, a releváns genetikai régióban ez az egyetlen olyan gén, aminek köze van a pigmentációhoz, ráadásul számos más háziszárnyasban bizonyították be, hogy EDNRB2 mutációi depigmentációt eredményeznek.
Ami azért is érdekes, mert úgy tűnik, hogy a – feltehetőleg – szabályozó mutáció hatása nem szorítkozik egyedül a szemre, és ugyanazok az egyedek, amelyek bull-típusú szemet hordoznak, testük számos egyéb területén depigmentáltak: a fejükön szinte mindig, de nagyon gyakran a szárnyukon és a nyakukon is.
Szóval itt állunk most, de még biztos, hogy ahogy a kutyák is szinte kimeríthetetlen forrásai a genetikai kutatásoknak, a galambokról is fogunk jópárszor hallani.
Domyan ET, Kronenberg Z, Infante CR, Vickrey AI, Stringham SA, et al. (2016) Molecular shifts in limb identity underlie development of feathered feet in two domestic avian species. eLife 5: e12115. doi: 10.7554/eLife.12115.
Boer EF, Maclary ET, Shapiro MD (2021) Complex genetic architecture of three-dimensional craniofacial shape variation in domestic pigeons. bioRxiv doi: 10.1101/2021.03.15.435516.
Maclary ET, Phillips B, Wauer R, Boer EF, Bruders R, et al. (2021) Two Genomic Loci Control Three Eye Colors in the Domestic Pigeon (Columba livia). bioRxiv doi: 10.1101/2021.03.11.434326.
Egy kellemes tavaszi délutánon az erdőszélen sétálva jó eséllyel botlunk lomha, nagy potrohú, fémesen csillogó kis drágakövekbe, amint hosszú, vézna lábaikkal esetlenül vonszolják testüket a talajon. Ők a hólyaghúzófélék családjába tartozó, és sokak által jól ismert nünükék (Meloe sp.). A nemzetségnek hazánkban hozzávetőleg egy tucatnyi faja él, és mindegyikükre jellemző, hogy lárvakorukban önjáró kakukkfiókaként anyjuknál sokkal fürgébb és gondoskodóbb nevelőszülőkre kényszerítik rá magukat. Egészen konkrétan méhekre.
A nőstény nünükék a peték lerakását követően röviddel elpusztulnak (ez plusz egy érv a nevelőszülők keresése mellett). A petékből tavasszal, vagy nyár elején kikelő háromkarmú (triungulinus) lárvák felkapaszkodnak a virágzó növényekre, és a porzószálak közt meglapulva várnak a gyanútlan beporzókra. Karmocskáikkal aztán az arra szálló méhek kitinszőreibe kapaszkodnak, és a fészekbe vitetik magukat. A célpontok jellemzően magányosan élő (szoliter) méhfajok, mint az Andrena, Anthophora és a Chalicodoma nemzetségek tagjai. A triungulinus lárva a méhsejtbe kerülve először az ott lévő tojást fogyasztja el, majd egy másik lárvastádiumot öltve a felhalmozott mézkészleteket kezdi felfalni. Mindeközben egy relatíve biztonságos helyen fejlődhet. Lenyűgöző és mindenekelőtt kényelmes életstratégia ez, a kleptoparazitizmus (más fajok zsákmányának, illetve önfenntartás vagy utódgondozás céljából felhalmozott készleteinek megdézsmálása) egy érdekes alesete.
Európai rokonainál egy fokkal erőszakosabb módon érnek célba egy amerikai nünükefaj, a Meloe franciscanus lárvái. Ez a faj az Egyesült Államok nyugati területeinek félsivatagos, sivatagos élőhelyein érzi magát igazán otthon. Egy-egy magasabb kórón a triungulinus lárvák egész kis tömege szokott összeverődni, egy magányos méhfaj nőstény egyedének sziluettjét és illatát utánozva. Ilyen magasszintű kooperációra euszociális rovarok körében bőséggel akad példa, de bogaraknál még csak hasonlót sem figyeltek meg eddig. Főleg egzotikus ez a viselkedési forma, mivel egy speciális mimikrivel is társul.
Egy szerelemtől vak, és párzásra kész hím méhet, ha sikerrel megtéveszt a Meloe franciscanus lárvák nyüzsgő kupacának látványa, és a lárvák által termelt feromonfelhő, amaz a párzás reményében túl közel merészkedik a lárvakupachoz, és ezzel kész is a baj. A lárvák megrohamozzák a méhet, és amennyien csak tudnak, belekapaszkodnak a bundájába. A testsúlyeloszlás hirtelen megváltozása miatt a lépre csalt méh gyakran drámai módon a földre zuhan, majd miután a lárvák kényelmesen elhelyezkednek rajta, és ő visszanyeri egyensúlyát tovább repül, hogy ezúttal egy valódi nőstényre akadjon. Miután méhünk sikeresen rátalált szíve választottjára, és beteljesült szerelmük, a nünükelárvák, mint valami nemi betegség, átragadnak a nőstényre. Innentől kezdve az ő bundáján laknak, és türelmesen várják, hogy a nőstény megépítse a fészket, lerakja petéit és megkezdje a méz felhalmozását. Ekkor aztán eljön a triungulinusok ideje! Belevetik magukat a mézbe, s innentől kezdve európai unokatestvéreikhez hasonló módon tobzódnak a jólétben.
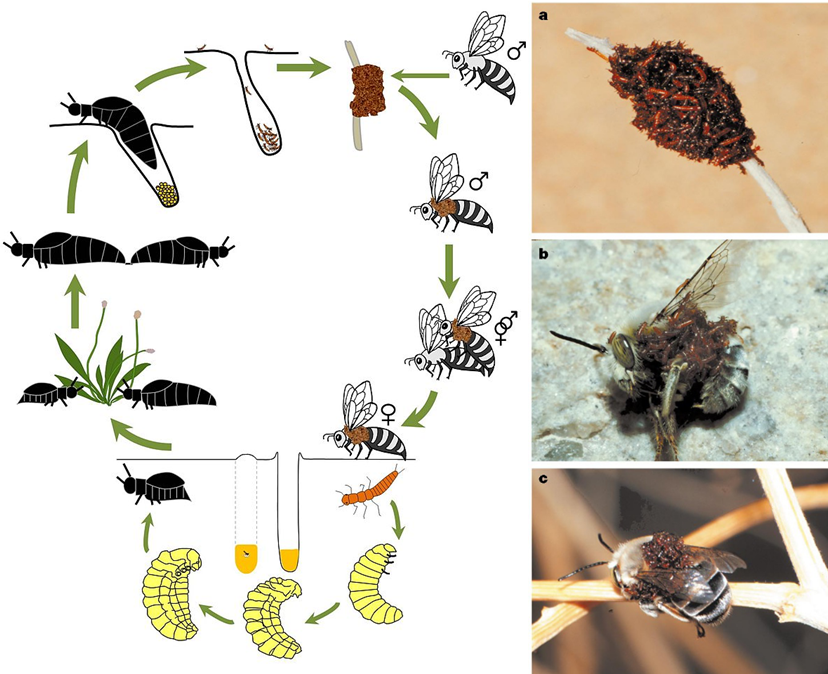
A Meloe franciscanus életciklusa (bal) (forrás: Wikipedia), triungulinus lárvák kupaca egy elszáradt növény szárán (a) és a megtévesztett méh testén (b; c) (forrás: Hafernik és Saul-Gershenz).
Belátható, hogy a M. franciscanus sikere azon áll, vagy bukik, hogy a lárvák magukhoz tudják-e vonzani a hím méhet. Minthogy ezt elsősorban az adott méhfaj nőstényeire jellemző feromonok termelésével érik el, a kleptoparazitizmus itt erősen gazdaspecifikus. A méhek ha szabadulni akarnak a nünükéktől, egyetlen evolúciós út járható számukra: változtatni kell egy kicsit a faj szexferomonjainak kémiai sajátosságain. Ez azonban könnyen reproduktív izolációhoz vezet az eredeti méhfajon belül, mivel az egyik populáció hímjei előbb utóbb nem lesznek képesek érzékelni egy másik populációba tartozó nőstény illatát. A populációk tehát előbb-utóbb faji szinten is elválnak egymástól.
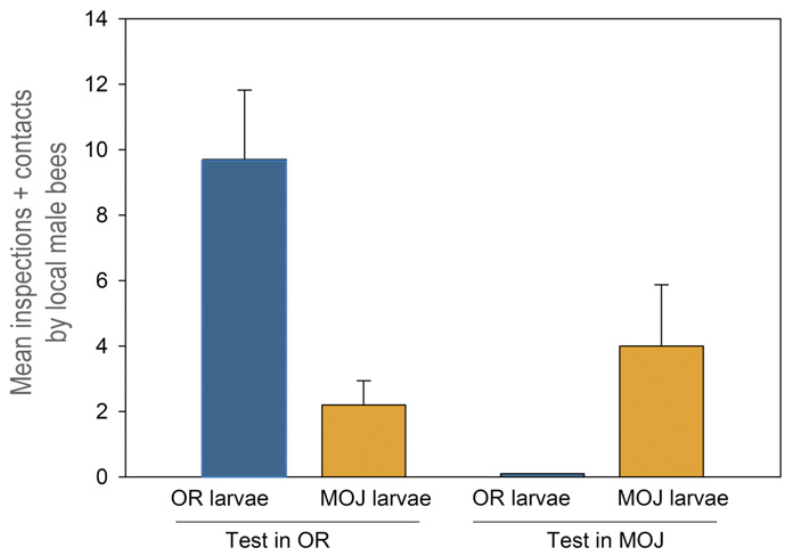
Persze a parazitának is lépést kell tartania a „feromonkódok” újraírásával, ha nem akar gazda nélkül maradni. Erre szépen rímel az a megfigyelés, hogy a M. franciscanus Oregon part menti dűnéin élő populációja a Habropoda miserabilis nevű oregoni méhfajra specializálódott, míg ugyan ennek a nünükének a Mojave-sivatagban élő populációja előbbi méhfaj egy itt élő, közeli rokonfaján, a Habropoda pallida-n élősködik. A nünüke és a méhek közti kémiai verseny tetten érhető, ha (miként azt Leslie Saul-Gershenz és munkatársai tették) elemezzük az általuk termelt feromonkoktélok összetételét.
A hártyásszárnyúak szexferomonjai többnyire páratlan, 21-31 szénatomból álló, egyszeresen telítetlen szénhidrogének (a kettős kötés leggyakrabban a 9. szénatomnál található, de a különböző fajok feromonkoktéljaiban ettől eltérő pozícióba is kerülhet). A két Habropoda faj feromonjai között jellemző különbségeket találtak, és fajon belül természetesen a hímek és a nőstények ferromonprofilja is egészen másképp fest. Szépen tükrözi viszont a gazda-parazita kapcsolatot jellemző kémiai kódíró versenyt, hogy az oregoni M. franciscanus feromonjai között több olyan vegyület is található, ami a H. miserabilis nőstényére jellemző, míg a mojavei nünükék, noha biztosan ugyan abba a fajba tartoznak, mint az oregoniak (ezt anatómiai és genetikai elemzések is igazolják), feromonprofiljuk jelentősen eltérő. A mojavei populáció feromonkoktélja több olyan komponenst tartalmaz, ami a H. pallida-ra jellemző.

A H. miserabilis és a H. pallida hímjeinek és nőstényeinek valamint az oregoni és Mojave-sivatagi nünükelárvák feromon profilja. A vízszintes tengelyen a különböző szénatomszámú feromonok vannak feltüntetve, kék színnel jelölve azokat, amelyek legalább két csoportban ugyan úgy jelen vannak. a függőleges tengelyen relatív koncentráció látható (Leslie Saul-Gershenz és mtsai, 2017).
A feromonprofil vizsgálata mellett olyan kísérleteket is végetek a szerzők, amelyek a két nünükepopuláció eltérő gazdaspecificitását igazolták. Ezekből kiderült, hogy a mojavei nünükepopuláció valóban csak az ott élő Habropoda fajt vonzza magához, míg az oregonit pedig csak alig, vagy egyáltalán nem, és vice versa. Ezekről a kísérletekről olvasva az ember elgondolkozik rajta, hogy az eltérő gazdaspecificitás a két nünükepopuláció egyedei között (a jelentős földrajzi távolságokat nem számolva) mennyire járul hozzá a populációk szétválásához, vagy adott esetben magához a fajképződéshez…
A jelen poszt forrásául szolgáló tanulmányok nem a tudomány legfrissebb termékei, és még csak azt se hiszem, hogy világmegváltó jelentőséggel bírnának. Amiért mégis beszámoltam róluk, annak csak az az oka, hogy idehaza épp „nünüke szezon” van. Bár a hazai Meloe fajok életciklusa kevéssé bizarr mint az M. franciscanus-é, ha a nyájas olvasó a hétvégén netán kirándulásra adná a fejét, és teszem azt egy kék nünükével (M. violaceus) találkozna gondoljon majd ezekre az érdekes amcsi rokonaira is 😉
Szép időt, és kellemes túrázást!
(A borítóképen egy kék nünüke látható – a szerző fotója)
__________
John Hafernik és Leslie Saul-Gershenz (Nature, Brief Communcations, 2000): „Beetle larvae cooperate to mimic bees„
Leslie Saul-Gershenz és mtsai. (PNAS, 2017): „Deceptive signals and behaviors of a cleptoparasitic beetle show local adaptation to different hostbee species„
Az első “teljes” neandervölgyi genom több mint egy évtizeddel ezelőtti publikálása óta tudjuk, hogy az anatómiailag modern ember (kvázi azt, amit Homo sapiens néven ismer a világ) Afrikából történő kivándorlása során keveredett az akkor már szélesebb körben elterjedt neandervölgyi populációkkal és ennek nyomát az Afrikán kívül élő emberi populációk genomjában mind fellelhtejük, nehány százaléknyi Homo neanderthalis DNS formájában.
Ennek a legősibb keveredésnek az idejét 50 – 60 000 évvel ezelőttre saccolják és feltételezett helye valahol a Közel-Keleten lehetett, ahol több nyomát is találták annak, hogy a két Homo faj párhuzamosan élt egymással. Később aztán az is egyértelművé vált, hogy a keveredés nem csak egyszeri esemény lehetett, hanem viszonylag rendszeresen bekövetkezett, erre volt jó példa a 2015-ben publikált, a romániai Csontok barlangjából (Peștera cu Oase) előkerült lelet, ahol a genomszekvencia alapján a 40 000 évvel ezelőttre datált személynek szépszülői magasságban lehetett neandervölgyi felmenője.
A héten a Nature-ben, illetve a Nature Ecology and Evolution-ben jelent meg egy cikkpáros, ami cseh és bulgáriai mintákat feldolgozva további (közvetett) bizonyítékkal szolgál arra vonatkozóan, hogy Délkelet-Európában viszonylag rendszeres lehetett az őshonosnak tekinthető H. neanderthalis és a több hullámban érkező H. sapiens-ek közti keveredés.
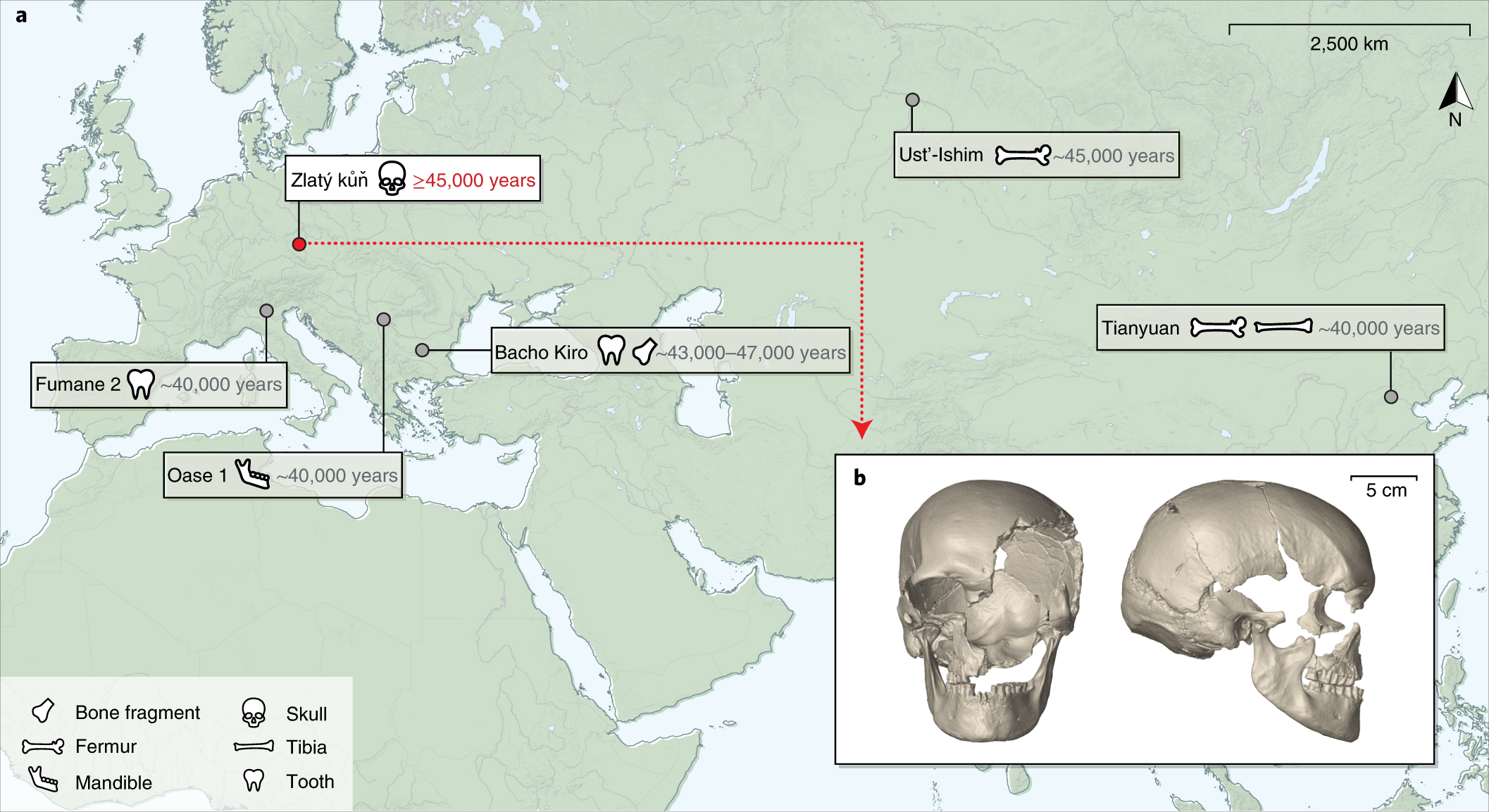
Előbbi közleményt a neandervölgyi genom feltárásban oroszlánrészt vállaló, illetve az Oase leletet is megszekvenáló Svante Pääbo csoportja jegyzi, és három olyan emberi maradványt vizsgáltak, amelyek a bulgáriai Bacso Kiro-barlangból kerültek elő (és egyelőre a legősibb leletek a térségben).
Az egy fogból és pár más csonttöredékből álló minta azonban így is felbecsülhetetlen értékű, mert egyrészt jól mutatja, hogy a térségben már felső- és középső paleolitikum határán folyamatos emberi jelenlét volt megfigyelhető (a barlang különböző rétegeiből előkerülő minták között van olyan, ami 45 ezer évvel ezelőtti és van olyan is, ami “csak” 35 ezer éves).
A régebbi minták esetében szintén viszonylag friss (kb. 7 generációval korábbról) származó neandervölgyi egyedekkel való keveredés nyomait hordozták. Vagyis a neandervölgyi anyag itt már csak kb. fele annyi volt, mint amit az Oase genomban láttak, de kicsit több, mint a fiatalabb bacso-kiroi mintákban találtak (utóbbiban a nendervölgyi szekvenciák aránya elég közel van ahhoz, amit a tipikus felső paleolitikumból származó mintákban találhatunk). A másik érdekesség, hogy míg az Oase genom esetében a genom modern emberi komponensének a modern emberi populációkban nem leljük nyomát (vagyis ez a csoport később kihalt), a bolgár leleteknek úgy tűnik van kapcsolata későbbi emberi populációkkal. Csakhogy ezek a kapcsolatok egyáltalán nem nyugat Eurázsiában vannak, ahogy az a minták földrajzi pozíciója alapján sejtenénk, hanem Kelet Ázsiában illetve Amerikai őslakói között.
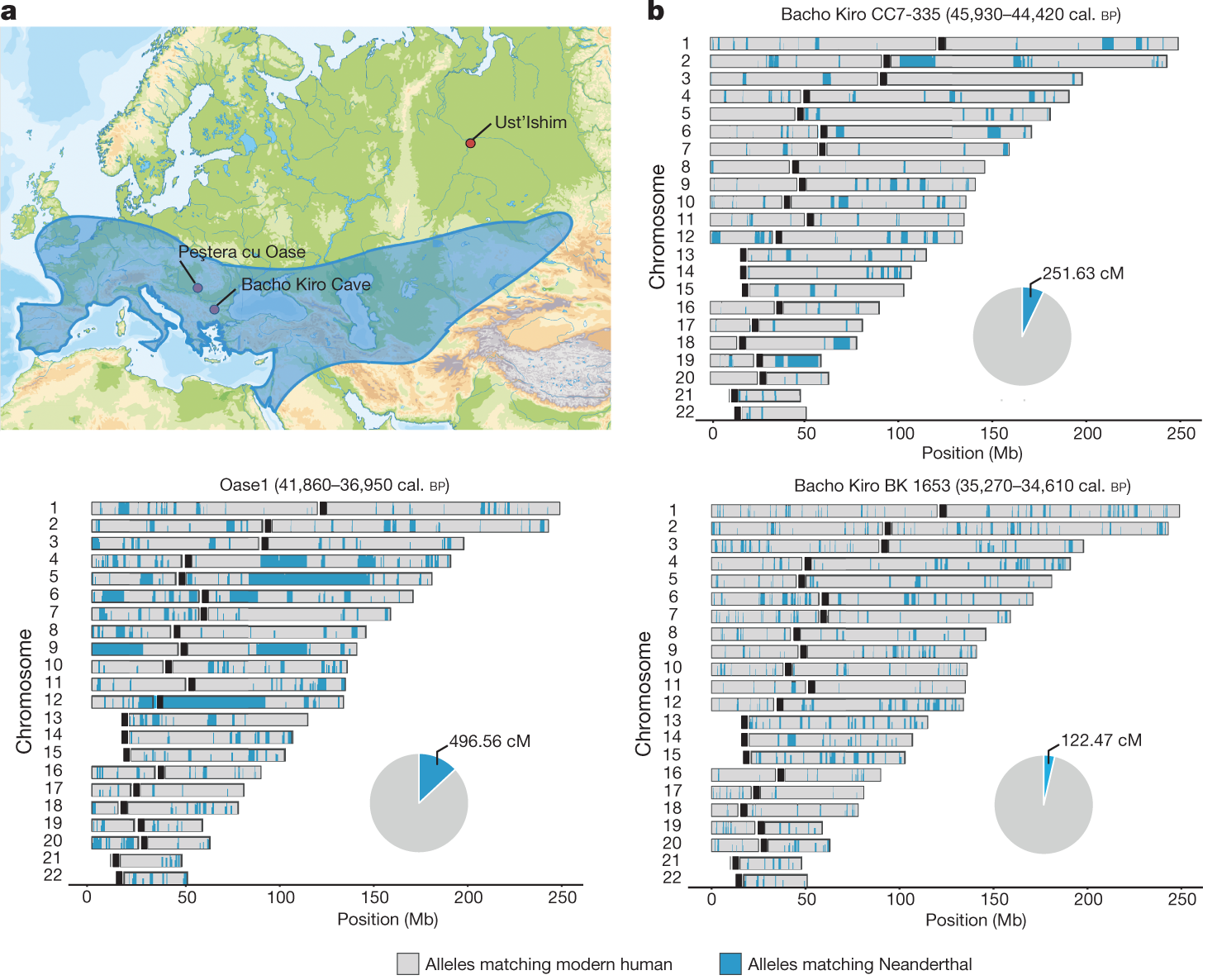
És ebből a szempontból megint más a cseh lelet, ami Zlatý kůň-ból kerül elő. Ez a több, mint 45 ezer évesre becsült, jó megtartású koponya, inkább a fiatalabb bulgáriai leletekhez hasonlít abból a szempontból, hogy a belőle kinyert genom neandervölgyi komponense nem kiemelkedő (bár kicsit több, mint a felső paleolitikumban talált mintákban), viszont az oasei lelethez kapcsolódik abból a szempontból, hogy nincs kontinuitása a ma is élő emberi populációkkal (a romániai lelettel való esetleges “rokonság” vizsgálata egyelőre várat magára).
Ez pedig egy érdekes kérdést vet fel: mi történt ezekkel a korai modern emberi populációkkal, amelyek már a középső paleolitikum végére láthatóan megtelepedtek a Kárpát-medencében, miért nem járultak hozzá a térségben később jelen levő vadászó-gyűjtögető populációkhoz?
Az egyik, talán leghihetőbb, magyarázat szerint egy vulkáni kitörés következtében kialakuló környezeti katasztrófa vezethetett a tréség ideiglenes elnéptelenedéséhez. Konkrétabban, a mai Nápolyhoz közel található Flegrei-mezők kialakulásában is kulcsszerepet játszó, 40 ezer évvel ezelőtt bekövetkező campaniai ignimbrit kitörés okozhatott olya környezeti változásokat, ami miatt a Kárpát-medence (is) ideiglenesen élhetetlenné vált.
Egy másik érdekesség azonban, ha már a H. sapiens és H. neanderthalis közti keveredésről beszélünk, az az, hogy ez egyelőre egyirányúnak tűnik. Vagyis azt látjuk, hogy a modern emberi populációkba bekerültek darabok a neandervölgyi genomból, de arra egyelőre nincs bizonyíték, hogy a késői és korai neandervölgyi genomok (és ha nem is sok, mostanra azért van pár) olyan szekvenciákban különböznének, amelyek H. sapiens eredetűek.
Ennek egyik magyarázata lehet, hogy nem áll még rendelkezésünkre elegendő neandervölgyi genom és a “mintavételezésünk” félrevezető, egyszerűen nem találtuk meg azokat az egyedeket, ahol ez a hibridizáció tetten érhető.
Egy másik magyarázat viszont, hogy mint olyan gyakran a hibridek esetében, az első generációs sapiens – neanderthalis hibridek csak az egyik szülői fajjal tudtak szaporodóképes utódokat létrehozni, részben a két faj között kialakuló genetikai inkompatibilitásnak köszönhetően. (Itt kb. arra kell gondolni, hogy egy komplexben együtt működő fehérjék egy fajon belül általában együtt változnak, de két nagyon régen elvált vonalban ellentétes irányban is változhatnak, így a hibridben már gondok lépnek fel a komplex működése során.) Ez nem új elmélet, már 2014-ben is komolyabban felvetődött, hogy a hibrid hímek sterilek lehettek azóta pedig több más eredmény is megerősíteni látszik.
Persze ezek a hibridizációk nem csak a középső paleolitikumban zajlottak, hanem ismételten előfordulhattak ahol és amikor csak az ősi H. sapiens és H. neanderthalis populációk egyazon területen éltek. És egy sokkal korábbi, (több) százezer évesre becsült hibridizációs esemény nyomát azért megtalálni a neandervölgyi genom két specifikus darabjában, a mitokondriális DNS-ben és az Y kromoszómában.
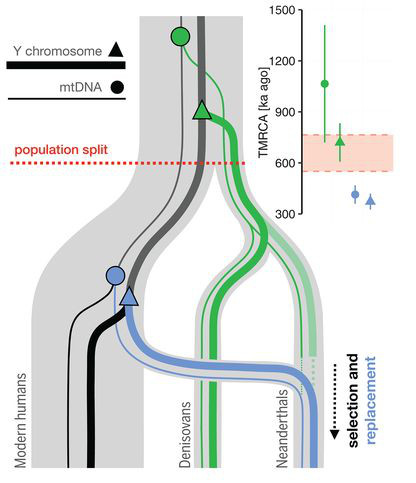
Ezek ugye genomunknak azoknak a darabjai, amelyek szigorúan anyai, illetve apai vonalon öröklődnek. Nem zavarja őket holmi rekombináció, ha esetleg összeszednek valami mutációt, akkor hűen tovább adják a lány, illetve fiú utódoknak.
A jelenleg ismert neandervölgyi genomok egyik érdekessége, hogy a belőlük izolált mitokondriális DNS (mtDNS), illetve Y kromoszóma alapján a neandervölgyiek közelebbi rokonok kellett volna legyenek a modern emberrel, mint a gyeniszovaival. Viszont a sokkal kiterjedtebb (sejtmagi) genom-adatok ezzel ellentétes képet festenek: eszerint a gyeniszovai és neandervölgyi populációk közös őse hamarabb vált el a modern ember ősétől.
Ez a két, látszólag egymásnak ellentmondó adat egy módon hozható közös nevezőre egy tavalyi Science cikk szerint: ha azt feltételezzük, hogy valamikor 100-370 ezer évvel ezelőtt az akkori ősi H. sapiens-ek és a neandervölgyiek közti hibridizáció eredményeképpen bekerült az ősi modern emberi mtDNS, illetve Y kromoszóma a neandervölgyiekbe és sikeresen “kiszorította” az ősi mtDNS és Y kromoszómás vonalakat. (Hogy ez esetben pontosan mit jelent a “kiszorította” arra max. ötletelhetünk, de ne feltétlenül gondoljunk egy aktív folyamatra, lehet hogy egyszerűen véletlen, hogy ezek az ősi “hibridek” maradtak fenn később Európában.) Ha ez valóban így van, akkor, ha valaha sikerülne mintát izolálni a spanyol Atapuerca közelében levő Sima de los Huesos barlangban talált kb. 400 ezer éves neandervölgyi mintákból, joggal remélhetnénk, hogy azok egy ősibb mtDNS-t mutatnának.
Végül, már inkább csak megjegyzés szintjén: a neandervölgyi introgressziónak jól dokumentálható hatása van ma is és ez lehet az egyik oka, hogy nem hígult ki teljesen a genomunkból. Például az immunrendszer működésében már korábban is találtak olyan neandervölgyi-eredetű génvariánsokat, amelyek erős szelekciós nyomás alatt vannak, így azt feltételezzük, hogy korabeli fertőzések esetében valamilyen előnyt adhattak hordozóinak. De, ezek a kis szekvenciák kétélű fegyverként is működhetnek.
Amikor a COVID-19 járvány elején felmerült, hogy a nagyon eltérő erősségű megbetegedéseknek genetikai oka is lehet, Pääbo és egyik munkatársa kimutatták, hogy az egyik DNS szakasz, amit kapcsolatba hoztak a súlyosabb lefolyású betegséggel, pont egy olyan genetikai variánshoz (allélhoz) kacsolható, ami – úgy tűnik – neandervölgyi eredetű.
Ez a harmadik kromoszómán levő régió az immunválaszban is fontos kemokineket (is) kódol. A betegségre hajlamosító allél (rs35044562 a pontos neve) hiányzik a modern afrikai genomokból, ellenben homozigóta formába van jelen az 50 ezer éves Horvátországból előkerült Vindija 33.19 neandervölgyi leletben, vagyis pont azt a mintázatot mutatja, amit klasszikusan a neandervölgyi genetikai introgresszióról gondolunk. Bár még a pontosan nem értjük, hogy az introgresszált régió miben különbözik, elképzelhető, hogy az erősebben aktivált immunválasz, ami más patogének ellenében hasznos lehet(ett), ebben járványban ellenkező hatást ér el.
(A borítókép forrása a Nature News volt.)
Hajdinjak, M., Mafessoni, F., Skov, L. et al. (2021) Initial Upper Palaeolithic humans in Europe had recent Neanderthal ancestry. Nature 592, 253–257. doi: 10.1038/s41586-021-03335-3.
Prüfer, K., Posth, C., Yu, H. et al. (2021) A genome sequence from a modern human skull over 45,000 years old from Zlatý kůň in Czechia. Nat Ecol Evol doi: 10.1038/s41559-021-01443-x.
Petr M, Hajdinjak M, Fu Q, Essel E, Rougier H, et al. (2020) The evolutionary history of Neanderthal and Denisovan Y chromosomes. Science 369: 1653-1656. doi: 10.1126/science.abb6460.
Zeberg, H., Pääbo, S. (2020) The major genetic risk factor for severe COVID-19 is inherited from Neanderthals. Nature 587, 610–612. doi: 10.1038/s41586-020-2818-3.
Még valamikor a 19. sz. végén, a kor egyik legjelesebb brit zoológusa, D’Arcy Thompson azzal szórakoztatta karácsonykor gyerekeit, hogy egy gumilapra kutyafigurát rajzolt és azt különböző irányba húzogatva egyik kutyafajtából csinált egy másikat. Thompson kvintesszenciális viktoriánus tudós volt és zoológia mellett nagyon érdekelte a matematika és fizika is (meg történetesen a klasszikus görög irodalom) és a gumihúzogatás a gyerekek szórakoztatásán túl egy mélyebb tudományos gondolat fizikai megtestesülése is volt. Utóbbi szerint az egyes élőlények kinézetét (a biológiai anyag formáját) elsősorban fizikai erők alakítják.
Ezt az elméletet öntötte aztán 57 évesen könyvformába “On Growth and Form” című művében, ami 1917-ben jelent meg és sokan máig a 20. század egyik legfontosabb biológiai témájú művének tartják. Pedig D’Arcy elsődleges szándéka, miszerint alternatívát kínáljon az öröklődés-alapú darwini evolúciós elméletnek, sosem kapott kellő támogatást. És mégis kellő mennyiségű érdekes gondolat került bele a műbe, hogy számos egymást követő kutatói generációt volt képes inspirálni. (Akit érdekelnek Thompson életének egyéb részletei, annak mindenképpen ajánlom Stephen Wolfram igazi fanboy-lelkesedéssel megírt hosszú írását.)

A karácsonyi gumikutyák logikája az “On Growth and Form” utolsó előtti fejezetében köszön aztán vissza, de itt (elsősorban) nem kutyákat, hanem halakat találunk. És ezek, a fent is látható rajzok lettek később a legikonikusabbak D’Arcy Thompson főművéből.
Mert bár az öröklődés fogja végső soron a testek alakját formálni, de azt nagyon jól ismerte fel Thompson, hogy bizonyos testrészek evolúciós átalakulása felfogható mint egy koordinátarendszer-torzuláshoz vezető növekedésbeli változás. Ezeknek pedig mind biológiai (genetikai) oka van, legyen az mondjuk a tacskók rövid lába, vagy a zsiráfok hosszú nyaka.
A halak esetében tényleg rengeteg ilyen típusú “transzformációt” figyelhetünk meg (persze nem teljesen véletlenül, hiszen a gerinces fajok fele az hal), és talán az egyik legérdekesebb közülük a repülőhalak mellúszójának esete.
Amiben egy tipikus repülőhal igazán különleges, az mellúszójának a mérete, amely egyben azt a relatíve nagy felületet is biztosítani tudja, ami ezeknek a fajoknak a siklórepüléséhez szükséges (és egy ilyen siklás akár 400 m hosszan is tarthat, miközben a magasságukat és a siklás irányát is szabályozzák). Hogy mi lehet ennek a megnagyobbodott mellúszónak az eredete, annak eredt a nyomába a Harvardon dolgozó Matthew Harris csoportja.
Az egyelőre csak preprint formájában elérhető kutatásban Harris-ék a repülőhalakat is magukba foglaló Beloniformes (magyarul Makrahalalakúak) rend 35 különböző fajának a fehérjekódoló, illetve konzervált, de nem kódoló szekvenciáját határozták meg. (Durva közelítéssel ezek a lényegi részei a genomnak, persze azért ez itt egy szükséges egyszerűsítés, ami ahhoz volt szükséges, hogy ennyi fajra költséghatékonyan meg lehessen oldani a szekvenálást.) Ezek segítségével meg lehetett becsülni, hogy milyen folyamatokhoz kapcsolódóak azok a fehérjekódoló gének, amelyek (feltehetőleg az adaptáció miatt) gyorsabban evolválódtak a repülőhalakban.
A gyorsan változó gének közt vannak olyanok, amelyek mondjuk az egyensúlyért felelős vesztibuláris rendszerhez kapcsolhatók, egyes izomfejlődésben fontos gének és számos olyan gén is, amelyet ilyen-olyan módon, más fajokban korábban már az uszony/végtag fejlődéséhez kapcsoltak. Ezek azok a gének, amelyeket jelen tudásunk alapján várnánk is, hogy változzanak, de persze egy ilyen összevetésben kijönnek olyan találatok is, amelyek általánosabb funkciót betöltő fehérjék és amelyek kevésbé “gyanúsak” egy ilyen esetben. Itt vannak például kálium-csatornákat (kcnk5a, kcnk9), vagy aminosav-transzportban fontos fehérjéket kódoló gének (lat4a, lat4b), amelyeket egy tipikus genomikai elemzésben még 4-5 évvel ezelőtt is a “futottak még” kategóriába soroltak. És ez az a pont ahol Harrisék csoportja szerencsére a genomikát egy jó adag (zebrahal)genetikával keverte össze, amitől igazán érdekes lett a történet.

Harris korábban a Nobel-díjas Christiane Nüsslein-Volhard egyik nagyon sikeres poszt-dokja volt és maga is részt vett abban a genetikai screen-ben, ahol olyan mutációkat kerestek, amelyek a felnőtt állatoknak változtatják meg valamiképpen a külalakját – mintázatát, pikkelyzetét, vázrendszerét, vagy épp az úszók méretét. Ez tipikusan olyan projekt volt, amihez egyrészt szükséges volt az a támogatás, amit egy Nobel díjas biztosítani tud, másrészt logikusan következett Nüsslein-Volhard (vagy, ahogy mindenki ismeri, Yani) korábbi munkáiból. Nobel díját Yani még azért a fantasztikus munkákért kapta, amit Eric Wieschaus-al együtt a korai muslica embriók fejlődésében szerepet játszó gének feltérképezése során végeztek (és ami az alapja minden fejlődésgenetika kurzusnak), majd ezután lényegében ugyanezt a munkát kiterjesztette zebrahal embriókra is. Ez volt ebben a fajban az egyik első komolyabb genetikai screen és egyben az egyik kulcsmomentum abból a szempontból is, hogy a George Streisinger által megálmodott modellszervezet valóban be tudott futni. De míg az embrionális genetikai screenek (főleg, ha muslicáról van szó), viszonylag gyorsak, addig ebben a legutóbbi kísérletsorozatban hónapokat, ha nem éveket kellett várni, amíg egyáltalán kiderült, hogy valóban új fenotípussal álunk szemben. Ebben a munkában játszott aztán fontos szerepet Harris.
Persze ekkor nem csak új mutánsokat kerestek, hanem a korábbi screenekből származó mutációkat is igyekeztek feltérképezni, így például azt is megérteni, hogy a nagyon eredetien longfin (lof) és another longfin (alf) névre keresztelt mutánsok, amelyek a nevüknek megfelelően meghosszabbodott úszókkal dicsekedhettek, milyen mutációknak köszönhetik létüket.
Az eredmény meglepő volt, hiszen ahogy először az alf esetében, majd később a lof-nál is kiderült, hogy a nagyobb úszóméretért kálium csatornák felelősek (előbbinél kcnk5b, utóbbinál pedig kcnh2a érintett). Mivel ezeket a csatornákat leginkább az idegrendszerben betöltött szerepük miatt tanulmányozták korábban, meglehetősen váratlan volt hogy megnövekedett expressziójuk eredményeképpen megnövekedett úszókat kaphatunk és egyben rávilágított, hogy a bioelektromos jelátvitel még sok más szövetben játszhat fontos szerepet.
Harris csoportja is a kcnh2alof mutációból indult ki (ők maguk is feltérképezték a mutáció helyét), hiszen ezek az úszók hasonlítanak leginkább a repülőhalak megnövekedett úszóira. Ugyanakkor a lof halakban a farokúszó mindkét része is megnyúlt ez pedig a repülőhalaknál nincs így. Ezért egy ellentétes hatású mutációt kerestek, egy olyant, aminek következtében rövidebbek lesznek az úszók és végül a nem túl fantáziadús nr21 “személyében” akadtak egy ilyen fenotípust okozó, domináns mutációra.
Az nr21 esetében a mutáció a lat4a génbe esett (itt is egy expressziót növelő, funkció-nyeréses mutációról van szó) és itt már érthető is, hogy miért ér össze a repülőhalak és a fura uszonyú zebrahal-mutánsok története, hiszen, ahogy fentebb írtam pont ilyen típusú gének mentek keresztól felgyorsult evolúción a repülőhalak genomjában.
Persze, ha van egy mutációnk, ami hosszabb úszókat hoz létre és egy másik, ami meg rövidebbet, adja magát a kérdés, hogy mi lesz, ha a kettőt összehozzuk? Harrisék megtették ezt és az eredmény pont azért izgalmas, mert kinézetében az így létrejövő kettős mutáns zebrahal (lat4anr21/+;kcnh2alof/+) eléggé hozza a repülőhalak “formáját”: elnyúlt mellúszókkal és felemás farokúszóval (felül rövid, alul hosszú) rendelkezik. Azaz két lépésben magunk is le tudtuk másolni a repülőhalak evolúciójának a talán leglátványosabb, thompsoni értelemben transzformációs lépését. (Természetesen mindez még nem elég a “repülőhalsághoz” és ezek a zebrahalak sosem fognak repülni. Ahhoz még számos más élettani és viselkedésbeli változásra lenne szükség.)
Bármennyire látványos is (és gumilapra kívánkozó) transzformáció, az “On Growth and Form”-ból sajnos pont a repülőhalak úszója hiányzik. De ez nem azért van, mert D’Arcy Thompsont esetleg nem érdekelték volna ezek a halak. A görög irodalomban nagyon is otthon levő Thompson maga fordította angolra Arisztotelész “Az állatok története” című művét, amiben a repülőhalak egyik első említése is található és ez kerülhetett be később, az “On Growth and Form” után három évtizeddel publikált “Glossary of Greek Fishes” című könyvbe is, amiben Thompson a klasszikus görög irodalomban említés szintén megjelenő halakat próbálta beazonosítani.
(A borítókép a Flickr-ről származik.)
Daane JM, Blum N, Lanni J, Boldt H, Iovine MK et al. (2021) Novel regulators of growth identified in the evolution of fin proportion in flying fish. bioRxiv 2021.03.05.434157; doi: 10.1101/2021.03.05.434157.
F.J. van Eeden, M. Granato, U. Schach, M. Brand, M. Furutani-Seiki, et al. (1996) Genetic analysis of fin formation in the zebrafish, Danio rerio. Development 123: 255-262.
Nüsslein-Volhard C. (2012) The zebrafish issue of Development. Development 139: 4099-4103 doi: 10.1242/dev.085217.
Perathoner S, Daane JM, Henrion U, Seebohm G, Higdon CW, Johnson SL, et al. (2014) Bioelectric Signaling Regulates Size in Zebrafish Fins. PLoS Genet 10(1): e1004080. doi: 10.1371/journal.pgen.1004080.
Stewart S, Le Bleu HK, Yette GA, Henner AL, Robbins AE, et al. (2021) longfin causes cis-ectopic expression of the kcnh2a ether-a-go-go K+ channel to autonomously prolong fin outgrowth. bioRxiv 790329; doi: 10.1101/790329.
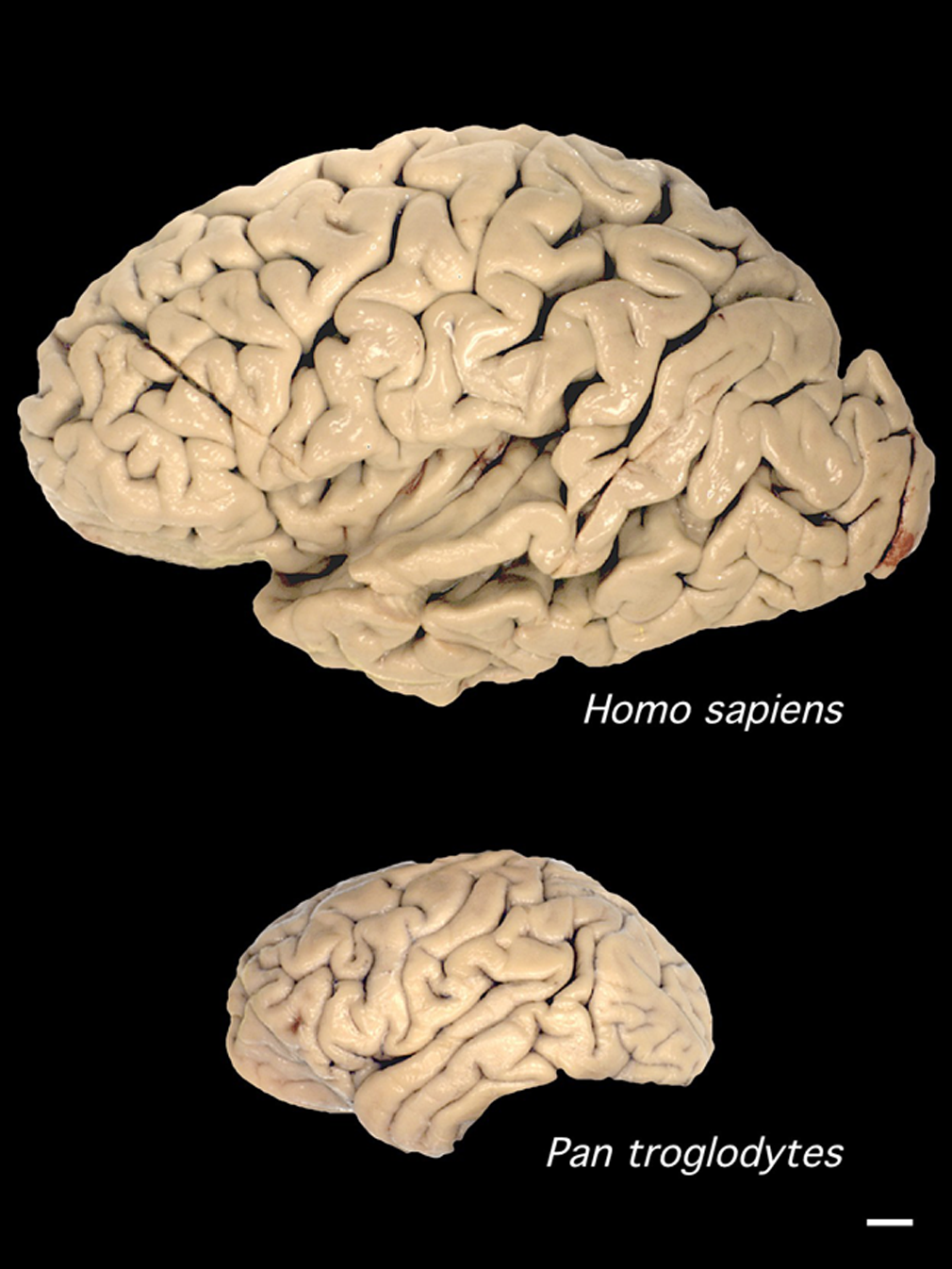 Bár a Fajok eredete igazából nem az ember eredetéről szól, mégis mindenki tudta és érezte már a megjelenésekor, hogy ez az az igazi kérdés, amit a benne leírt tudományos elmélet feszeget. Darwin, a rá jellemző visszafogottsággal, ebben a könyvében csak egy rövidke utalást tesz a kérdésre (“Fény derül majd az ember eredetére és történetére. (Light will be thrown on the origin of man and his history)”, amolyan beharangozóként későbbi, másik főművéhez, ami Az ember származása és az ivari kiválás lenne. Mindez jól tükrözi, hogy már bő másfél évszázada (ha pont nem “már a görögök óta”) az egyik legfontosabb kérdés, ami sok biológiai kutatás középpontjában is áll, arra keresi a választ, hogy mi az, ami minket fajként különlegessé tesz és megkülönböztet más fajoktól.
Bár a Fajok eredete igazából nem az ember eredetéről szól, mégis mindenki tudta és érezte már a megjelenésekor, hogy ez az az igazi kérdés, amit a benne leírt tudományos elmélet feszeget. Darwin, a rá jellemző visszafogottsággal, ebben a könyvében csak egy rövidke utalást tesz a kérdésre (“Fény derül majd az ember eredetére és történetére. (Light will be thrown on the origin of man and his history)”, amolyan beharangozóként későbbi, másik főművéhez, ami Az ember származása és az ivari kiválás lenne. Mindez jól tükrözi, hogy már bő másfél évszázada (ha pont nem “már a görögök óta”) az egyik legfontosabb kérdés, ami sok biológiai kutatás középpontjában is áll, arra keresi a választ, hogy mi az, ami minket fajként különlegessé tesz és megkülönböztet más fajoktól.
Itt a blogban már közel másfél évtizede is írtam egy hosszabb posztot arról, hogy mi különböztet meg minket főemlős rokonaiktól és amint abban is látszik, természetesen a kulcskérdés szinte mindig az agyra vonatkozik. Hiszen vannak egyéb érdekes vonásaink (pl. relatív szőrtelenségünk), de mégis az agyunk, pontosabban az agyunk által “kódolt” szellemi képességeink azok, amelyek minden jel szerint különlegessé tesznek bennünket – nem természetfelettivé, de mégis egy olyan fajjá, ami a saját eredetén és képességein gondolkodni tud.
Így aztán minden kor a maga technológiai repertoárjának megfelelően próbált nekiállni az agy vizsgálatának. Amíg csak morfológiát lehetett nézni, addig azt nézték előszeretettel és hasonlították más emlősök, főként emberszabásúak agyának morfológiájához. Amikor később mikroszkópos szintű vizsgálatokra kerülhetett sor, sokan a különböző neuronok morfológiájának, számának és kapcsolatainak feltárásában próbálták keresni a különbségeket. És amikor már genomszekvenciák is elérhetővé váltak, akkor vagy az egyes csimpánz/gorilla/ember szekvencia-szakaszok összevetését vették előszeretettel elő, vagy mindent megtettek, hogy az esetleges különbözőségek biológiai értelmét megfejtsék in vivo kísérletekkel.
Természetesen, mivel emberen és főemlősökön (nagyon helyesen) nem igazán lehet kísérletezni, korábbi technológiák esetében az in vivo kísérlet elsősorban azt jelentette hogy különböző gén-szakaszok emberi és csimpánz megfelelőit fejeztették ki egérben.
Az elmúlt években az in vitro , vagyis “Petri-csészében” nevelt, jellegzetes struktúrát és differenciációs mintázatot mutató 3D-s sejttenyészetek, az organoidok és még konkrétabban az agy organoidok (populárisabb nevükön “miniagyak”) formájában egy teljesen új technológia vált elérhetővé. Ezeknek a miniagyak használata egyébként is érdekes és fontos etikai kérdéseket vet fel, és mivel alapvetően érdekes dolgokat árulhatnak el az idegrendszer korai fejlődéséről, nyilvánvalóan csak idő kérdése volt, hogy valaki felhasználja őket a köztünk és más emberszabásúak közti különbségek boncolgatására.
Ez a “valaki” aztán annak a Madeline Lancasternek a csoportja lett, aki Jürgen Knoblich bécsi laborjában poszt-dokként eredetileg is az úttörője volt az agy organoidok készítésének és tanulmányozásának.
Az új Cell-ben közölt tanulmányban csimpánz és gorilla “miniagyakat” vetettek össze a (most már rutinból készített) emberi agy organoidokkal. Külsőre ezek eléggé hasonló struktúrákat hoztak létre, azzal a különbséggel, hogy az emberi organoidok jellemzőek nagyobbak voltak, mint a másik két emberszabásúé, bennük egy viszonylag nagyobb üreggel.
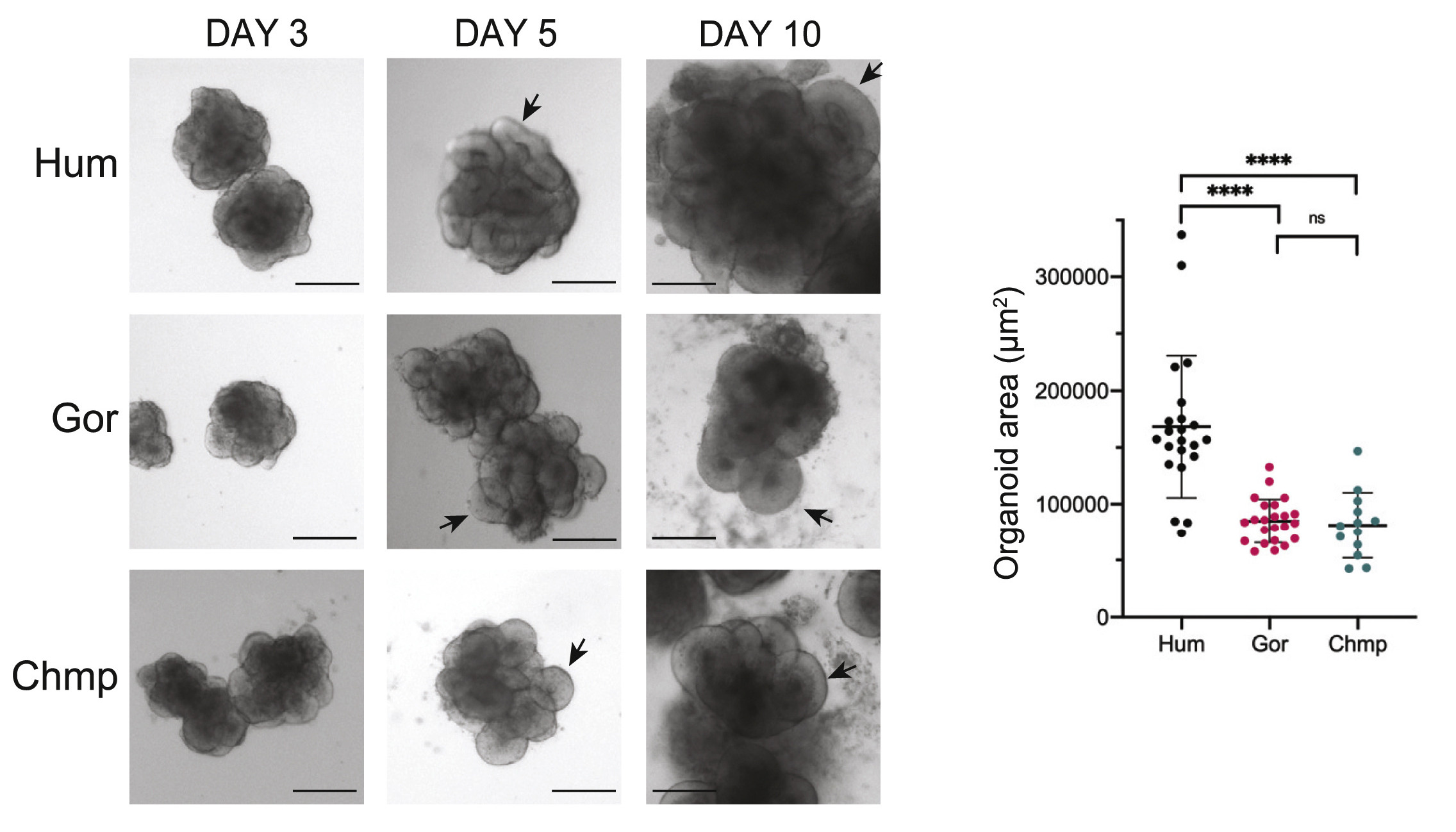
Mindez arra utalt, hogy az emberi organoidokban egységnyi idő alatt több sejt keletkezik. Persze ezt igazolni is kellett, és végül több módszer segítségével ezt meg is tették. Ezeknek a megfigyeléseknek az érdekese hozadéka az a megfigyelés, hogy az emberi organoidokban az idegrendszeri progenitor sejtek gyorsabban osztódnak és később kezdenek el differenciálódni.
Az embrionális fejlődés során is az idegszövet kezdeti, “éretlen” állapota egy olyan ún. neuroepitélium (NE), ahol kicsit elnyúlt sejteket találunk, amelyek a kialakuló szövet mindkét oldalával (apikális, illetve bazális, szakzsargonban) kapcsolatban van. A sejtmagok egy jellegzetes jojó-mozgást végeznek ebben a stádiumban, oda-vissza mozognak a két felszín közt a sejtciklusuk alatt (ún. interkinetikus nukleáris migráció) és az apikális felszín közelében osztódnak.
Az organoidokban is ez ismétlődik meg (az apikális felszín az organoid belsejében nyíló üreg oldalán lesz) és az emberi miniagyakban gyorsabb ez a mozgás és későbbre tolódik az a pillanat, amikor a NE sejtek elkezdenek átalakulni (tNE), ami az első jele annak, hogy megkezdődik az idegszövet differenciációja.

2021-ben nincs már magára valamit is adó cikk rettenetesen sok szekvenálási adat nélkül, így itt is rengeteg stádiumban meghatározták a fejlődő emberi és gorilla miniagyak génexpresszióját, olyan géneket keresve, amelyek egy kicsit más viselkedést mutatnak a két rendszerben.
A kísérlet logikája az volt, hogy feltételezhető, a kicsit eltérő sejtciklusok mögött valami szabályozó génnek a megváltozott expressziója állhat – ami, tegyük hozzá, azért egy nem minden kockázat nélküli hipotézis, főleg, hogy az ilyen típusú “adathorgász” megközelítések rengeteg potenciális lehetőséget szoktak feldobni, amelyek közül nagyon sok egyszerűen a szekvenálási adatokban jelen levő biológiai “zaj” eredménye és nincs igazi biológiai relevanciája. Végül azonban találtak/kiválasztottak egy ZEB2 nevű gént, amit tovább is vizsgáltak.
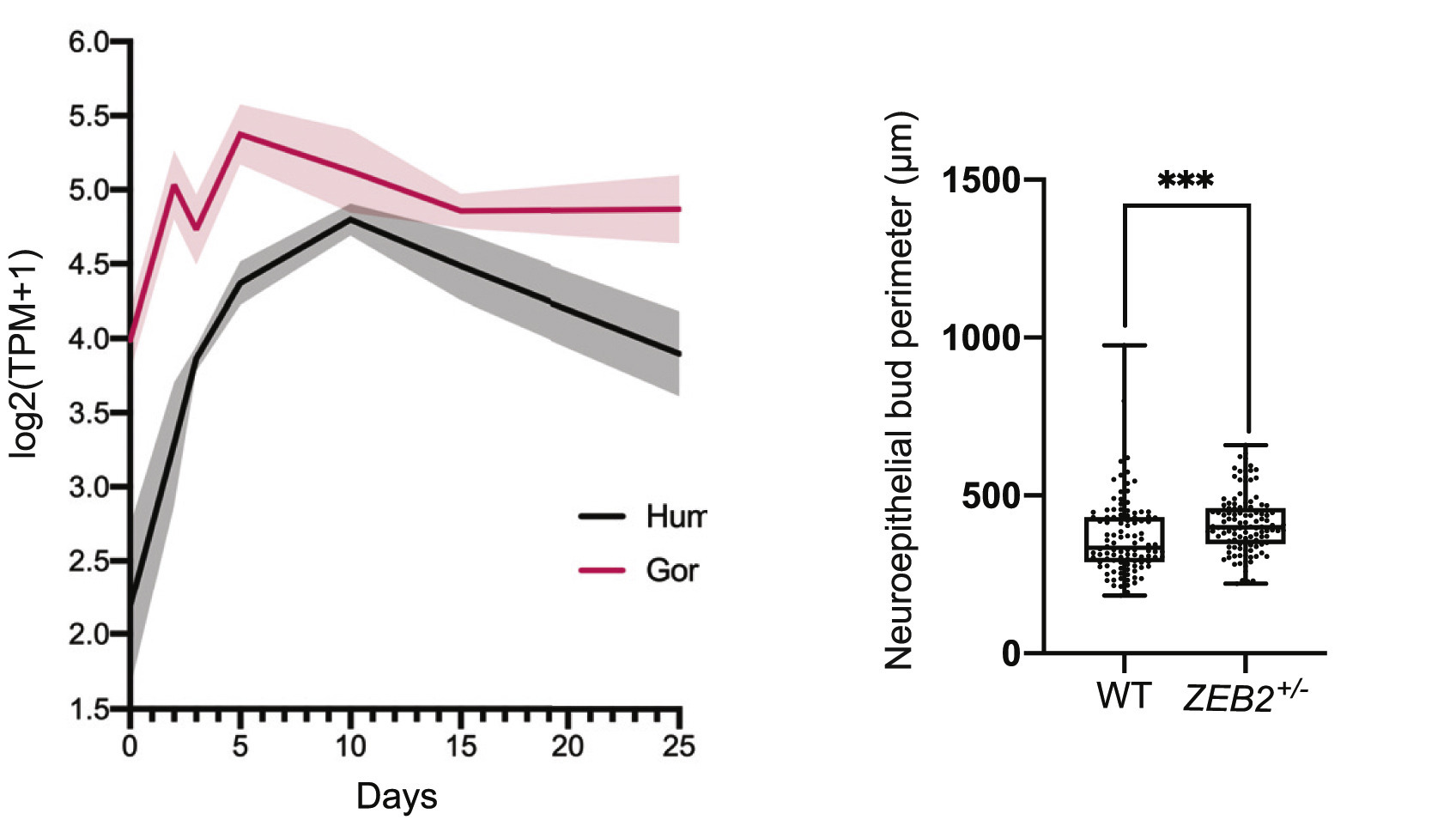
Azért a ZEB2-re esett a választásuk, mert egyrészt ez pont olyan expressziós profillal rendelkezik, hogy egy picit hamarabb van az expressziós csúcsa a gorilla organoidokban, mint az emberben, másrészt maga a gén is számos olyan jelet mutat, hogy az ember felé vezető leszármazási vonalban felgyorsult az evolúciója és pár nagyon jellegezetes aminosav-változást hordoz.
Amikor aztán olyan emberi miniagyakat hoztak létre, amelyekben lecsökkentették a ZEB2 funkcióját, azok kicsit, de azért mérhetően nagyobbak lettek. Hogy ez olyan szignifikáns változás, ami valóban egyértelműen bizonyítja a ZEB2 evolúciós szerepét valószínűleg vitatható lesz: míg a három faj organoidjai közt egyértelmű méretkülönbségek voltak, a ZEB2-hiányos organoid esetében a Lancaster-csoport már csak az organoidok oldalán megjelenő kis kitüremkedések (bud perimether) tudtak valamit kimérni.
A ZEB2 fehérje egyébként BMP-jelátviteli útvonal modulátoraként léphet itt fel és azt szabályozhatja, hogy mennyire hamar kezdődik el az a morfológiai átalakulás az apikális oldalán a NE sejteknek, ami aztán a differenciáció (vagyis a sejtciklusból való kilépés) felét tett egyik első fontos lépés lehet.
Összességében a cikk sok szempontból fontos mérföldkő lehet az ilyen típusú kutatásokban, de ahogy a korábbi hasonló tanulmányok esetében, még azért el kell egy kis időnek telnie, hogy meg tudjuk ítélni, mennyivel kerültünk valóban közelebb az emberi agy különlegességének megértéséhez.
Benito-Kwiecinski S., Giandomenico SL, Sutcliffe M, Riis ES, Freire-Pritchett P, et al. (2021) An early cell shape transition drives evolutionary expansion of the human forebrain. Cell https://doi.org/10.1016/j.cell.2021.02.050.
;_Fragment_of_a_Letter_or_Document_(verso)_MET_DP800417.jpg){kind=link}