
Még a blog hajnalán futó, “Csökevényszervek” sorozatban írtam egyszer az emberi farokról, ami egyrészt azért érdekes, mert jobbára nincs, pedig az emberi fajt is magába fogadó rendszertani törzs, a Gerinchúrosok egyik legfontosabb bélyege az ún. posztanális farok, másérszt – ahogy az ominózus poszt is pedzegeti – néha, atavisztikus jegyként mégis feltűnik.
A teljes képhez még hozzá kell tennünk, hogy mindez igaz más emberszabásúakra is, illetve a farok tulajdonképpen elkezd kifejlődni embrionálisan, de aztán, jobbára a sejtek pusztulása (apoptózisa) miatt, az utolsó trimeszterben elsorvad.
Mindig is érdekes kérdésnek tűnt, hogy vajon mi okozhatta evolúciósan egy ilyen kulcsbélyeg eltűnését, miért volt erre szükség, illetve hogyan figyelhető meg mégis pár emberben csökevényes farok a születés után. Ezekből az első és utolsó kérdésre ad, ha nem is teljes, de részleges választ egy napokban a bioRxiv-on publikált preprint.
A szerzők fontos meglátása volt, hogy az emberi gerinc és farok fejlődésének egyik kulcsgénje, a TBXT esetében az emberszabásúak genomja egy fontos pontban eltér más főemlősökétől: a hatodik és hetedik exonok közt feltűnik egy új ugrálógén-kópia, az AluY.
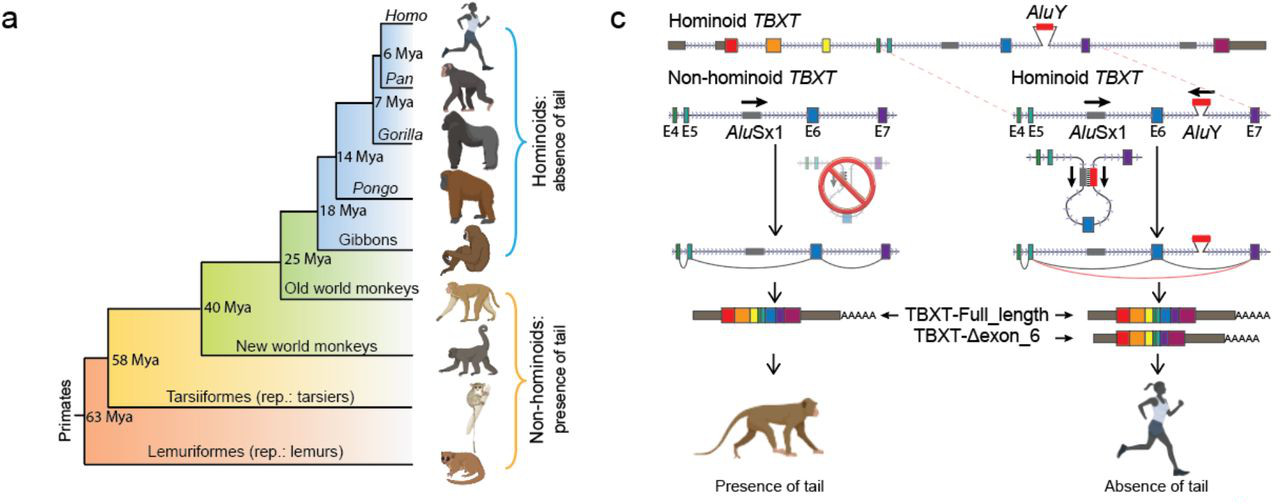
Az Alu elemek alkotják az emberi (pontosabban a főemlős) genom egyik legnagyobb “ugrálógén” közösségét, hiszen olyan, egyenként kb. 300 bp hosszú transzpozonokról van szó, amelyekből közel 1 millió van az emberi genomban, vagyis a genomunk több mint tizede ilyen szekvenciákból áll. Nem egy monolitikus transzpozontengerről van szó, ezeken belül is megkülönböztetünk különböző alcsaládokat, amelyek közül pár, így például a talán legfiatalabbnak tartott AluY is, még mindig aktívnak tűnik.
És ez adja a konkrét megfigyelés lényegét is, ugyanis a főemlősök genomjában a TBXT gén hatodik intronjában egy (evolúciósan) új AluY található meg. Ez azért vált különösen érdekessé, mert egy kicsit távolabbi rokona egy AluSx1 transzpozon az előző, vagyis ötödik intronban is felfedezhető és a két elem szekvenciahasonlósága kellően nagy, hogy a TBXT gén átíródásakor a még éretlen pre-mRNS-ben egy hurokszerű struktúra jöjjön létre, ami bezavar a pre-mRNS érésébe, azaz a splicing folyamatába.
Így emberben (és feltehetőleg más emberszabásúakban is) tulajdonképpen a TBXT gén két izoformája jelenik meg: a teljes hosszúságú és egy másik, amiből a hatodik exon hiányzik és egy csonkolt fehérje képződését segíti elő. Vagyis egy emberszabású embrió kis túlzással úgy viselkedik, mint egy heterozigóta mutáns (ahol szintén két verziója lenne jelen a génnek), s innen jött az ötlet, hogy mi lenne, ha meg is néznénk pont ezt egy olyan állatban, ahol egyébként kifejlődne a farok, pl. egy egérben.
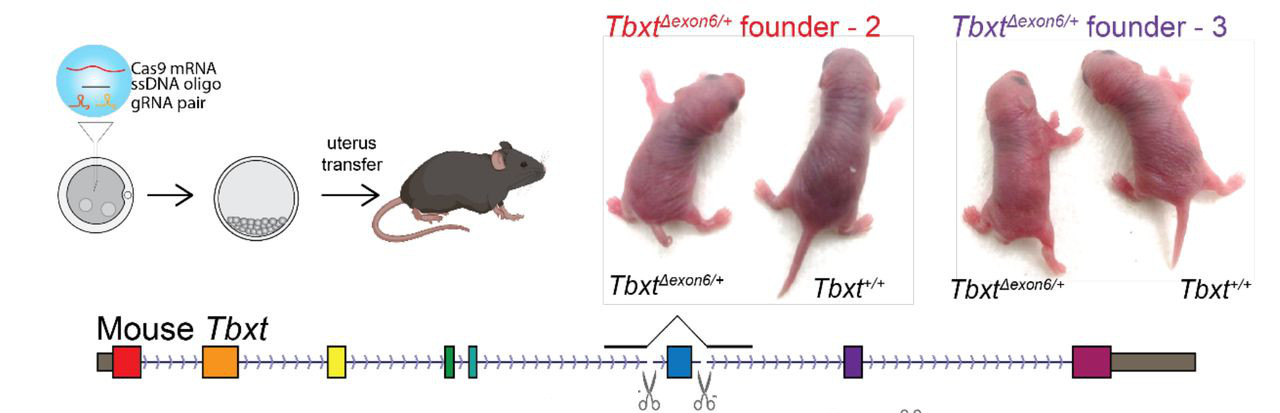
Modern genomszerkesztési technológiákat felhasználva létre is hoztak ilyen egereket és bingó, azoknak többnyire ilyen-olyan farok-fejlődési rendellenessége volt, többeknek pedig egyáltalán nem fejlődött ki a farka (összesen a hetrozigóták harmadánál volt valamilyen fenotípus). Ezeket a kísérleteket még kiegészítették emberi sejtvonalakon végzettek is, amelyek szintén visszaigazolták, amit sejteni lehetett: mind az AluY, mind az AluSx1 szükséges a plusz izoforma kialakulásához, hiszen egyik, vagy másik hiányában nem alakul ki a pre-mRNS érésbe bezavaró hurok-struktúra.
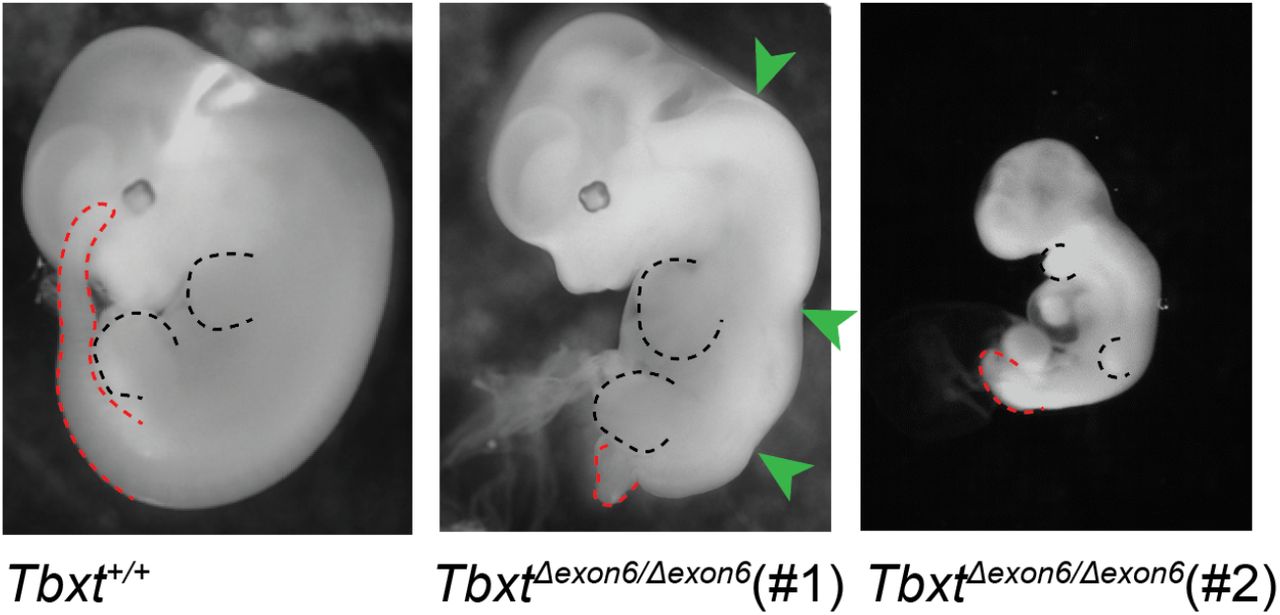
A mutáns egereknek köszönhetően azt is tudjuk, hogy a hatodik exon deléciója homozigóta állapotban halálos és az ilyen egerek a farok hiánya mellett az emberi nyitott gerincre (spina bifida) emlékeztető elváltozásokat mutattak.
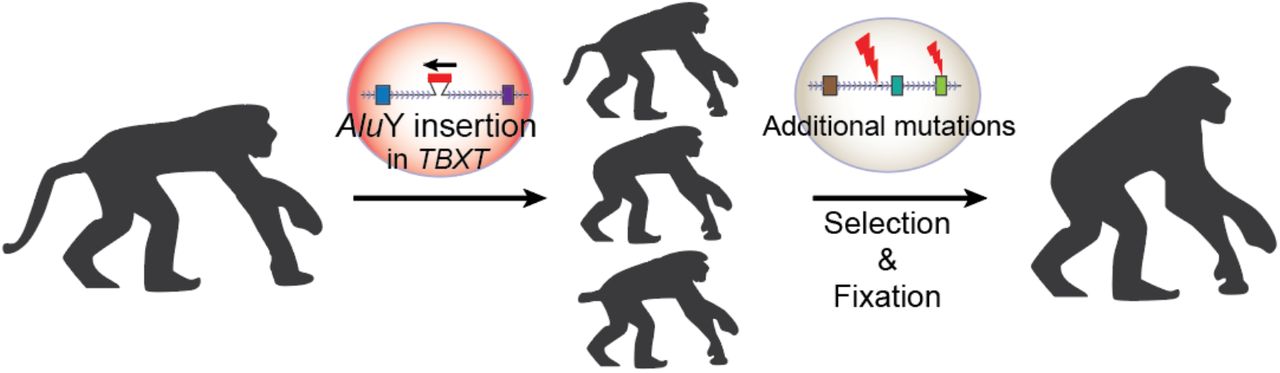
Összességében tehát úgy tűnik, hogy az emberszabásúak faroktalanságát egy transzpozon túlbuzgóságának köszönhetjük, amelynek következtében a TBXT gén két izoformája is jelen van a sejtekben. Valamiért (s itt azért még tapogatózunk) a faroknélküliség egy előnyt jelenthetett a földön több időt töltő őseinknek és megjelenhettek és rögzülhettek olyan stabilizáló mutációk is, amelyek a csak részben penetráns faroktalan fenotípust (a heterozigóta egerek csak harmada lesz az), teljesen penetránssá tették. Ugyanakkor, ha valamilyen egyéb okokból változik a két TBXT izoforma aránya, jó eséllyel annak vagy az lesz a következménye, hogy nyitott gerincű gyerekek születnek, vagy épp olyanok, akiken ektopikus, atavisztikus farkak jelennek meg.
(A fedőkép a Wikimedia oldaláról származik.)
;_Fragment_of_a_Letter_or_Document_(verso)_MET_DP800417.jpg){kind=link}
Xia B, Zhang W, Wudzinska A, Huang E, Brosh R et al. (2021) The genetic basis of tail-loss evolution in humans and apes. bioRxiv 2021.09.14.460388; doi: 10.1101/2021.09.14.460388