Néhány hete már esett szó arról, hogy a di- és trinukleotid
ismétlődéseknek (mikroszatellitáknak) milyen kulcsszerepe lehet az
evolúciós folyamatokban. Az akkori postban utóbbiakra kerestem példát,
bemutatva, hogy milyen hatása lehet annak, ha ezen ismétlődések száma a
fehérjét kódoló DNS szekvencián belül változik. Azonban számos fajban a
genomot alkotó DNS nagy része nem kódol fehérjéket, így logikus, hogy
számos mikroszatellita növekedése vagy csökkenése nem érinti
közvetlenül a kódoló régiókat. Érintheti ellenben az egyes gének
szabályozó részeit, mint hamarosan kiderül.
Logikus okokból ahhoz, hogy a genomban megjelenő kismértékű
változások és génexpressziós vagy viselkedésbeli különbségek között
egyértelmű megfeleltetést találhassunk, arra van szükség, hogy a
vizsgált egyedek közötti genetikai különbségeket minimalizáljuk, azaz
ezek lehetőség szerint azonos vagy közeli rokon fajokhoz tartozzanak.
Jó példa erre, hogy míg egyes amerikai egér (Microtus) fajok
(pl. prériegér) életük végéig tartó monogám kapcsolatban élnek, addig
közeli rokonaikra (hegyi egerek) ez egyáltalán nem jellemző, sőt
előbbiekkel szöges ellentétben az apák egyáltalán nem vesznek részt az
utódgondozásban. Azt már korai vizsgálatok kiderítették, hogy a két faj
között az egyik különbség a vazopresszin-1a receptor (v1ar)
expressziójában rejlik, s ennek egyik lehetséges oka a szabályozó
régióban található mikroszatellita hosszában felfedezhető eltérés lehet
(a szociális fajokban hosszabb, míg az antiszociális fajokban
rövidebb). Mivel azonban a vizsgált fajok között természetesen más
genetikai eltérések is voltak, nem lehetett kijelenteni, hogy a
viselkedésbeli különbség teljesen egyértelműen ehhez a dinukleotida
ismétlődéshez köthető-e.
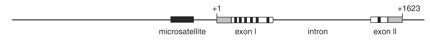
1. Ábra: A v1ar gén szabályozó szekvenciájában egy mikroszatellita található.
A kutatók segítségére sietett azonban a természetes variabilitás,
hiszen, mint kiderült, a prériegerek különböző földrajzi populációi
között is megfigyelhetők különbségek a V1aR szabályozó szekvenciájának
szóban forgó részében, még ha nem is annyira drasztikusak, mint az
egyes fajok között. S így ennek segítségével természetesen ki lehetett
szűrni a genetikai „háttérzajt” és azt vizsgálni, hogy milyen hatása
van önmagában a mikroszatellita méretének a gén kifejeződési területére
és az egerek viselkedésére.
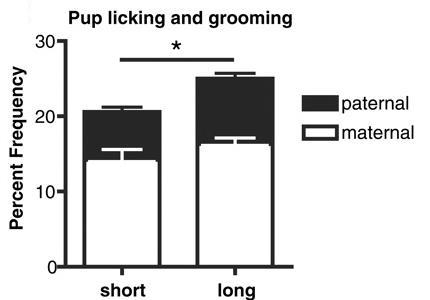
2. Ábra: A hosszabb dinukelotida ismétlődést hordozó hím egerek gondosabbak az utódgondozásban, mint rövid ismétlődésű társaik.
Mint
az kiderült, a hosszabb ismétlődéssel rendelkezo hímek sokkal
szociálisabbak és szignifikánsan több időt töltenek mind párjukkal,
mind az utódok pátyolgatásával, mint rövid ismétlődéssel rendelkező
rokonaik (2. ábra). Sőt, a V1aR expressziója is különbözik az egyes
példányok között (3. ábra), az egyik fontos különbség pont a rágcsálók
viselkedése szempontjából fontos szaglógumóban figyelheto meg.
Ezen adatok miatt különösen érdekes lehet, hogy az emberi V1aR gén
szabályozó részében négy mikroszatellita is található, amelyek közül
ketto egy deléció következtében hiányzik a csimpánzokból, de jelen van
a párkapcsolatuk szorosságáról ismert bonobókban. Sőt, néhány
megfigyelés szerint valamiféle összefüggés létezhet az autizmus és az
említett ismétlődések között. Természetesen óvatosan kell bánni a
nagyívű következtetések levonásával, de érdekes további vizsgálatok
tárgya lehet, hogy mennyiben befolyásolja a V1aR szabályozása az emberi
interakciókat.
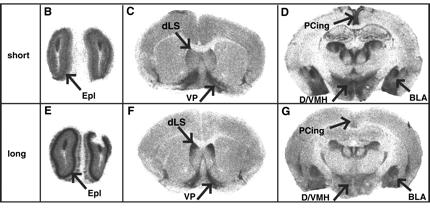
3. Ábra: A mikroszatellita hossz és a génexpresszió között erős
összefüggés van. A hosszabb szekvencia intenzívebb expressziót okoz a
szaglógumóban (B vs E) és a dorzolaterális septumban (dLS) (C vs F),
két olyan agyi területen, melyeket korábbi megfigyelések a
viselkedéshez kapcsoltak. Ugyanakkor egyes kortikális területeken és az
amygdalában csökken az expresszió (D vs G).
Hammock EAD and Young LJ (2005) Microsatellite instability generates diversity in brain and sociobehavioural traits. Science 308: 1630-1634.