
Luca napja van, legyen hát szó Lucáról, pontosabban LUCA-ról, a ma
élő organizmusok utolsó közös őséről (Last Universal Common Ancestor).
Nem tudjuk pontosan, hogy hogyan is nézhetett ki LUCA, de bizonyos
sejtéseink vannak néhány tulajdonságáról. Először is mivel a genetikai
kód univerzális és számos olyan fehérje ismert amelyek orthológjai
elővilág mindhárom nagy ágában (Archea, Eubacteria, Eukarya)
fellelhetők, meglehetős nagy valószínűséggel kijelenthetjük, hogy ezen
tulajodnságok egyben LUCA tulajdonságai is voltak. Az elterjedtebb
elméletek szerint LUCA leginkább egyes ma élő Gram-pozitív
baktériumokra hasonlíthatott, azaz bakteriális kör alakú DNS
kromoszómával, sejtmembrán és -fal által körülvett transzkripciós és
transzlációs apparatussal rendelkező prokarióta lehetett. Vagy nem,
legalább is ezt sugallja Eugene Koonin és William Martin egy
közelmúltban megjelent cikkjükben.
A Trends in Genetics véleményrovatában közölt írásukban arra
hívják fel a figyelmet, hogy az említett hasonlóságok ellenére számos
alapvető különbség is akad az Archea és Eubacteria csoportba tartozó
mikroorganizmusok között. A legismertebb talán az, hogy sejtmembránjuk
(és ami azt illeti sejtfaluké is) összetétele alapvetően különbözik: az
előbbiét izoprén-éterek, míg utóbbiét zsírsav-észterek alkotják. Sőt a
DNS másolásra is igen eltérő szekvenciájú fehérjéket használnak. Ennek
egyik magyarázata természetesen lehet az, hogy mivel az Archea
csoportba tartozó mikróbák elsöprő többsége extremofil, azaz
szélsőséges körülmények (magas hő és só koncentráció, vagy igen hideg)
között éldegél, az új életmódjukhoz való alkalmazkodás során
következtek be ezek a változások. Koonin és Martin azonban egy másik
lehetséges megoldást ajánl: LUCA még nem rendelkezett sem
sejtmembránnal, sem DNS alapú genommal, eze a két ágban később
egymástól függetlenül alalkultak ki. Igen ám, de a természetesen
felemrülő kérdés az, hogy akkor (szerintük) hogyan jöhetett létre
egyáltalán LUCA, hiszen lehetetlen, hogy egy akár ennyire komplex
organizmus spontán szerveződjön az „őslevesben”.
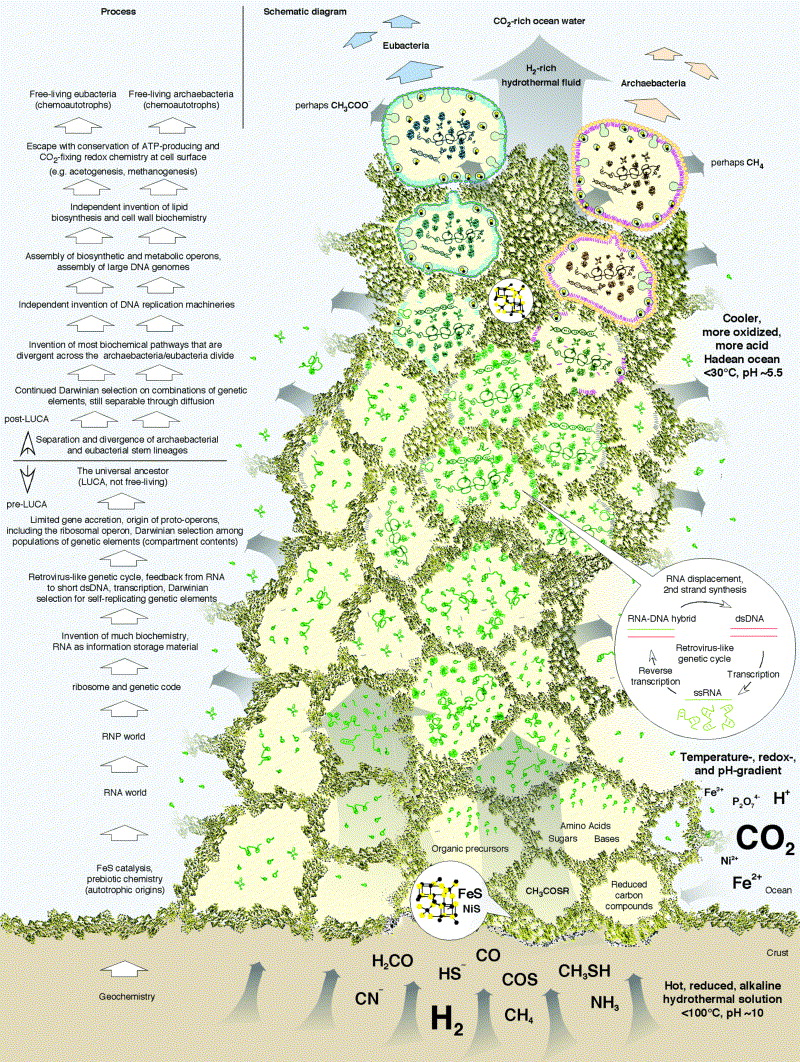
A két kutató által vázolt lehetőség sokban építkezik a viszonylag
közismert tengerfenéki hőforrás elméletre (e szerint az élet
keletkezése ezekhez a képződményekhez kötött). Szerintük LUCA egy ilyen
hőforrás környékén, geokémiai erők által létrehozott lyukacsos
kőzetekben jelenhetett meg. Az ilyen geológiai képződmények sok kb
1-100 mikron átmérőjű hasadékot tartalmaznak, és a forrás aktivitás
alatt ill. miatt állandóan növekednek, azaz állandóan új rekeszek
adódnak a képződményhez. A szintén jelen levő vas-szulfid katalizátora
lehetett a különböző biomolekulák létrejöttének, amelyek ezáltal egy
abiotikus, de kisméretű térben „lelték magukat”. A folyamat fő
hajtóereje (ezúttal is) az önreplikáció irányába mutató természetes
szelekció, ami egyrészt az előbb több, majd egyetlen szegmensen
(szakaszon) kódolt altruista operonok megjelenéséhez vezetett (amelyek
magukban kódolták a replikációhoz szükséges első géneket), másrészt
lehetővé tette az első paraziták megjelenését is (akik a már említett
replikációs apparátust kihasználták saját sokszorozódásukra, de ők nem
kódoltak a folyamathoz szükséges fehérjét). És ezzel kb. elértünk az
RNS-világhoz.
Mivel az egyes apró hasadékok között léteztek nyílások, az egyes
kompartmentekből időnként egy-egy molekula populáció egyszerű diffúzió
útján átkerült egy-egy szomszédos rekeszbe. Ez természetesen szelekciós
tényező is volt, hiszen amelyik társaság nem tartalmazta/kódolta a
replikációhoz szükséges összes elemet, az rövid időn belül nyomtalanul
eltűnt. A fejlődés egy következő pontja lehetett, amikor ez az ősi
genom egyfajta retrovírus-szerű replikációra tért át, azaz az RNS genom
replikációjába közbe iktatódott egy kettősszálú DNS is, amiről egy RNáz
segítségével íródott vissza a megfelelő orientációjú RNS (ezzel lehetne
magyarázni pl. az RNáz-H jelenlétét mind az Archea-ban, mind az
Eubacteria-ban).
És ez az a pont ahol az Archeák és Eubacteriák története elválik, azaz
az előbb vázolt molekulaegyüttes nem más mint LUCA. Az önreplikációra
képes primitív genomok ezután különböző úton, egymástól függetlenül
tértek át a teljesen DNS alapú genomra (ezért különböznek a DNS
replikáció komponensei annyira), ill. alakítottak ki maguknak
sejtmembránt. (A felmerülő probléma itt csak annyi, hogy a sok
különbözőség ellenére, van azért néhány membránfehérje is amelyik
mindkét csoportban megtalálható. Szerzőink szerint ezt nem lehet
későbbi horizontális gén transzferrel magyarázni – amely egyébként nem
ritka a prokarióták között -, ugyanis az ominózus fehérjék alapján
készülő filogenetikai fák egyértelműen elválasztják az Archea és
Eubacteria csoportokat. Az általuk elképzelhetőnek tartott magyarázat
szerint ezek a fehérjék az abiotikus hasadékok falán spontán képződő
hidrofób rétegekben létezhettek, s mivel ezek képződéséhez nincs
szükség egy membrán-szintetizáló apparátusra, nem feltételezi azt, hogy
LUCA genomja ilyesmit kódolt volna. Lehet, de szvsz ez az elmélet egyik igen gyenge pontja.)
Nagyításhoz katt a képre
Ez mind szép és jó, de ahhoz, hogy tudományos elméletként
fogadhassuk el kell legyen valami predikció, mert különben csak
filozófia. Nos, ezt természetesen a szerzők is belátják, ezért a
következő logikus potenciális falszifikációs utat javasolják:
amennyiben a sikerül olyan prokariótákra lelni, melyeknek Eubacteriára
jellemző membránjuk, de Archae típusú DNS szintetizáló apparátusuk van, az
teóriájuk instant halálát jelentené, hiszen annak értelmében ilyenfajta
„átmeneti” fajok nem létezhetnek. Mindenesetre ha valaha kiderül, hogy
elméletük igaz (ami egy elég nagy HA), az a szóban forgó egykori
mélytengeri hőforrást, ahol LUCA megjelent, valóban az „élet-forrásává”
tenné….
Koonin EV, Martin W. (2005) On the origin of genomes and cells within inorganic compartments. Trends Genet 21(12): 647-54. Epub 2005 Oct 11.