 Az autó előtt volt a ló, s ez időben a fehér hátas számított az abszolút státuszszimbólumnak. Nem meglepő hát, hogy a tenyésztők mindent megtettek a privilegizált kevesek ezirányú igényeinek kielégítéséért. E törekvések eredményeként, egymástól függetlenül, több világos lóparipafajta jött létre, melyek többnyire különböző mutációknak köszönhetik színüket. Az egyik ilyen genetikai elváltozásról nemrég már ejtettünk szót, most lássunk a „fehérló-probléma” egy másik lehetséges „megoldását”.
Az autó előtt volt a ló, s ez időben a fehér hátas számított az abszolút státuszszimbólumnak. Nem meglepő hát, hogy a tenyésztők mindent megtettek a privilegizált kevesek ezirányú igényeinek kielégítéséért. E törekvések eredményeként, egymástól függetlenül, több világos lóparipafajta jött létre, melyek többnyire különböző mutációknak köszönhetik színüket. Az egyik ilyen genetikai elváltozásról nemrég már ejtettünk szót, most lássunk a „fehérló-probléma” egy másik lehetséges „megoldását”.
Pontosabban, ha teljesen hűek akarunk maradni a valósághoz, ezek a lovak nem is fehérek, a szó szoros értelmében. Legalábbis életük elején nem azok (hanem szürkék), csak viszonylag hamar „megőszülnek”, s szőrük fehérré változik. Ennek a pontos biológiai (sejt szintű) mechanizmusa még nem ismert, de a korai őszülés mögött levő gén(ek) azonosításával, egy igencsak jó képet alkothatunk arról, mi is történik.
Azt már eddig is tudni lehetett, hogy a tulajdonság 25. kromoszómához kötődő, domináns jelleg (vagyis elég ha csak a kromoszómapár egyik tagján megtalálható a mutáció, akkor is látható lesz a hatása). Ami csak most derült ki, hogy pontosan, hol is van a kormoszómán a genetikai változás.
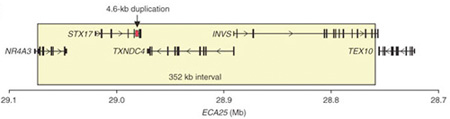 Több mint 700 ló megvizsgálása után (na és persze a jól haladó ló genom-projectnek is köszönhetően) a kutatást végzők egy szűkebb régióra kezdtek összpontosítani, ahol mindössze négy gén található. Ezek egyikét sem hozták korábban összefüggésbe a színezettel, bár sejtbeli funkciója alapján legalább az egyikük esetében nem zárható ki elviekben ilyesmi. Ez pedig egy synatxin gén (STX17), amelynek a sejt belsejében levő kis „csomagok”, ún. vezikulumok szállításáért felel: mivel a színanyagot termelő melanocita sejtek ilyen csomagokban tartják a színanyagukat, már látszólag meg is van a tettes. A gond, elsőre, csak az, hogy a STX17 fehérje szerkezetileg teljesen normális (sőt, a másik három fehérje esetében is hasonló a helyzet). Ha azonban a fehérjét kódoló gént vesszük jobban szemügyre, akkor hamar felfedezhető, hogy egyik nem kódoló régiója, intronja megduplázódott. S mivel ebben a DNS szakaszban olyan szabályozó elemek vannak, amelyek a gén expresszióját alapvetően meghatározzák, a duplikáció eredményeképpen a STX17 gén túltermelődik.
Több mint 700 ló megvizsgálása után (na és persze a jól haladó ló genom-projectnek is köszönhetően) a kutatást végzők egy szűkebb régióra kezdtek összpontosítani, ahol mindössze négy gén található. Ezek egyikét sem hozták korábban összefüggésbe a színezettel, bár sejtbeli funkciója alapján legalább az egyikük esetében nem zárható ki elviekben ilyesmi. Ez pedig egy synatxin gén (STX17), amelynek a sejt belsejében levő kis „csomagok”, ún. vezikulumok szállításáért felel: mivel a színanyagot termelő melanocita sejtek ilyen csomagokban tartják a színanyagukat, már látszólag meg is van a tettes. A gond, elsőre, csak az, hogy a STX17 fehérje szerkezetileg teljesen normális (sőt, a másik három fehérje esetében is hasonló a helyzet). Ha azonban a fehérjét kódoló gént vesszük jobban szemügyre, akkor hamar felfedezhető, hogy egyik nem kódoló régiója, intronja megduplázódott. S mivel ebben a DNS szakaszban olyan szabályozó elemek vannak, amelyek a gén expresszióját alapvetően meghatározzák, a duplikáció eredményeképpen a STX17 gén túltermelődik.
A már-már idilli képet azonban egy makacs tény nem hagyja összeállni: ha ezekben a lovakban valóban a kis „csomagocskák” szállításával lenne elsősorban gond a szőrzetért felelős szőrtüszők melanocitáiban, akkor az már születésüktől fogva evidens kellene legyen. De nem az, hiszen amint írtam születésükkor még igencsak színesek. Azaz mindent összevetve valószínűtlen, hogy a STX17 lenne a ludas. De akkor mi más?
Mint kiderült, a duplikáció nemcsak a synatxin gén expresszióját változtatta meg, hanem a szomszédos NR4A3 génét is megnövelte. Ennek szerepe a sejtciklus szabályozásában van, így például azt is meghatározza, hogy milyen ütemben jöjjenek létre a már emlegetett melanociták az őssejtjeikből. Ha túl sok NR4A3 van a sejtben a ciklus túlpörög és a túlerőltetett őssejtek elpusztulnak, vagyis egy idő után már nem jönnek létre új melanociták a szőrtüszőben, s ennek következményeként nem termelődik új színanyag. Ezt a „felpörgetős” modellt látszik alátámasztani az is, hogy a fiatal lovak, az őszülés beállta előtt, paradox módon kicsit besötétülnek – épp mint azt elvárnánk, ha kezdetben, az őssejtek kifáradása előtt, túlpörög a színanyag termelés.
A sejtciklusba való belepiszkálásnak azonban másfajta hatásai is lesznek: a 15 évnél idősebb szürke lovakban 70-80 százalékos valószínűséggel bőrrák lesz diagnosztizálható. Ennek elsődleges oka a melanociták egy másik típusának, a bőrben levő pigmentsejteknek a kóros elburjánzása. Különösen magas a dagant kialakulásának a gyakorisága a lipicai lovakban, így érdemes volt annak is utánanézni, hogy ezt milyen (további) mutáció okozhtaja?
A STX17 intron-duplikáció mellett ezekben a hátasokban az agouti nevű gén működésével is gondok vannak. Ez elsőre nagyon furcsa: más fajokban, ha ez a gén nem működik, annak sötét szőrzet az eredménye. A lipicai lovakban azonban a szőrtüszők melanocitáinak hiányában minderre nincs lehetősége az agouti által befolyásolt MC1R-jelátviteli-útvonalnak. Helyette viszont a bőrben levő további pigment sejtekben kavarnak (az STX17-el karöltve), s ennek lesz eredménye a melanoma.
(A lipicai lovak képe a kalandtabor.hu-ról származik.)
Rosengren Pielberg G, Golovko A, Sundström E, Curik I, Lennartsson J, et al. (2008) A cis-acting regulatory mutation causes premature hair graying and susceptibility to melanoma in the horse. Nat Genet 40(8): 1004-1009.

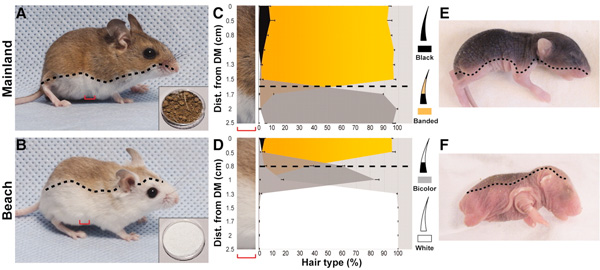
 Méltatlanul elhanyagoltuk egyik kedvenc témánkat, a
Méltatlanul elhanyagoltuk egyik kedvenc témánkat, a  És akkor egy kis visszatérés egyik kedvenc vesszőparipánkhoz, a pigmentáció evolúciójához. Ezúttal az észak amerikai szürkefarkasokat vesszük nagyító alá, ugyanis ennél a fajnál egy érdekes színezeti trend figyelhető meg: az északi, fátlan tundrán fiatalon világosszürkék, idősebben pedig piszkosfehér színűek ezek a ragadozók, viszont délebbre, ahogy nő az erdővel borított területek aránya, úgy nő egy olyan színváltozat gyakorisága is, amelyik fiatalon fekete, majd később sötétszürkévé válik.
És akkor egy kis visszatérés egyik kedvenc vesszőparipánkhoz, a pigmentáció evolúciójához. Ezúttal az észak amerikai szürkefarkasokat vesszük nagyító alá, ugyanis ennél a fajnál egy érdekes színezeti trend figyelhető meg: az északi, fátlan tundrán fiatalon világosszürkék, idősebben pedig piszkosfehér színűek ezek a ragadozók, viszont délebbre, ahogy nő az erdővel borított területek aránya, úgy nő egy olyan színváltozat gyakorisága is, amelyik fiatalon fekete, majd később sötétszürkévé válik.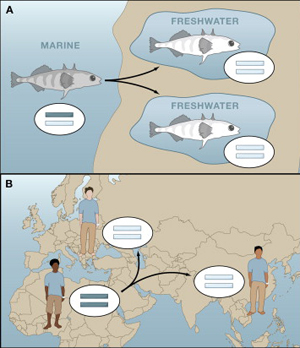 Az elmúlt év legvisszatérőbb témája az állati szőrszínezet, vagyis pigmentáció evolúciója volt, így talán stílszerű, hogy az év utolsó postjának tervezett, de persze az új évbe átcsúszó beírás is ezzel a témával foglalkozzon.
Az elmúlt év legvisszatérőbb témája az állati szőrszínezet, vagyis pigmentáció evolúciója volt, így talán stílszerű, hogy az év utolsó postjának tervezett, de persze az új évbe átcsúszó beírás is ezzel a témával foglalkozzon. A fehér ló regék és mítoszok visszatérő szereplője, mesebeli királyfik és valódi kormányzók választott hátasa. Pedig a fehér ló is csak ló, egyetlen különlegessége színében rejlik.
A fehér ló regék és mítoszok visszatérő szereplője, mesebeli királyfik és valódi kormányzók választott hátasa. Pedig a fehér ló is csak ló, egyetlen különlegessége színében rejlik. Hát akkor ismét szőrszín, mert ez (szinte) kifogyhatatlan téma, már csak azért is, mert a laikusok számára is evidens, könnyen érzékelhető változásról van szó.
Hát akkor ismét szőrszín, mert ez (szinte) kifogyhatatlan téma, már csak azért is, mert a laikusok számára is evidens, könnyen érzékelhető változásról van szó.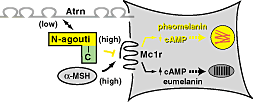 Megnyugtató módon, már a legelején tisztázódott, hogy valami úton-módon, a β-defensin is a klasszikus, Mc1r függő jelátviteli útvonalba szól bele. Sőt, kitartóbb vizsgálódás azt is bizonyította, hogy a fehérje fekete szőrszínt okozó formája erősen tud magához a receptorhoz kötődni, aktivizálva a fekete pigment szintézisét katalizáló folyamatokat. Mindez szép, logikus és kerek, de van egy bökkenő: a β-defensin gén „normál” (nem fekete színt okozó) verziója is képes hasonló erősséggel a receptorhoz kötődni, azaz a kapcsolat puszta ténye önmagában nem lehet a válasz.
Megnyugtató módon, már a legelején tisztázódott, hogy valami úton-módon, a β-defensin is a klasszikus, Mc1r függő jelátviteli útvonalba szól bele. Sőt, kitartóbb vizsgálódás azt is bizonyította, hogy a fehérje fekete szőrszínt okozó formája erősen tud magához a receptorhoz kötődni, aktivizálva a fekete pigment szintézisét katalizáló folyamatokat. Mindez szép, logikus és kerek, de van egy bökkenő: a β-defensin gén „normál” (nem fekete színt okozó) verziója is képes hasonló erősséggel a receptorhoz kötődni, azaz a kapcsolat puszta ténye önmagában nem lehet a válasz. Úgy tűnik, hogy hirtelen nagyon trendi lett neandervölgyi géneket vizsgálni, hiszen
Úgy tűnik, hogy hirtelen nagyon trendi lett neandervölgyi géneket vizsgálni, hiszen 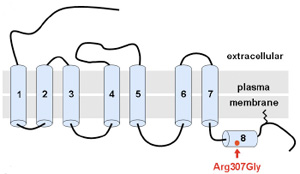 A neandervölgyi mc1r különlegessége, hogy bizonyos esetekben a jelek szerint egy olyan jellegzetes mutációt hordoz(ott), aminek a következtében, működésbe lépésekor a cAMP koncentráció nem éri el azt a szintet, amit a sötét hajú emberekben figyelhetünk meg. Érdekes módon, ez nem azért van (volt), mert a receptor kevésbé hatékony, hanem mert (pontosan nem tisztázott okok miatt) eleve kevesebb mutáns receptorfehérje jut el a sejtek belsejéből a felszínükre. A végeredmény szempontjából azonban ez mindegy és a létrejövő alacsonyabb cAMP szint megegyezik azzal, amit vöröshajú emberekből izolált MC1R hoz létre – azaz jó eséllyel feltételezhető, hogy az ősi neandervölgyiek kb. 1%-a vörös fej- és arcszőrzettel büszkélkedhetett.
A neandervölgyi mc1r különlegessége, hogy bizonyos esetekben a jelek szerint egy olyan jellegzetes mutációt hordoz(ott), aminek a következtében, működésbe lépésekor a cAMP koncentráció nem éri el azt a szintet, amit a sötét hajú emberekben figyelhetünk meg. Érdekes módon, ez nem azért van (volt), mert a receptor kevésbé hatékony, hanem mert (pontosan nem tisztázott okok miatt) eleve kevesebb mutáns receptorfehérje jut el a sejtek belsejéből a felszínükre. A végeredmény szempontjából azonban ez mindegy és a létrejövő alacsonyabb cAMP szint megegyezik azzal, amit vöröshajú emberekből izolált MC1R hoz létre – azaz jó eséllyel feltételezhető, hogy az ősi neandervölgyiek kb. 1%-a vörös fej- és arcszőrzettel büszkélkedhetett. A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.
A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.