Egy kellemes tavaszi délutánon az erdőszélen sétálva jó eséllyel botlunk lomha, nagy potrohú, fémesen csillogó kis drágakövekbe, amint hosszú, vézna lábaikkal esetlenül vonszolják testüket a talajon. Ők a hólyaghúzófélék családjába tartozó, és sokak által jól ismert nünükék (Meloe sp.). A nemzetségnek hazánkban hozzávetőleg egy tucatnyi faja él, és mindegyikükre jellemző, hogy lárvakorukban önjáró kakukkfiókaként anyjuknál sokkal fürgébb és gondoskodóbb nevelőszülőkre kényszerítik rá magukat. Egészen konkrétan méhekre.
A nőstény nünükék a peték lerakását követően röviddel elpusztulnak (ez plusz egy érv a nevelőszülők keresése mellett). A petékből tavasszal, vagy nyár elején kikelő háromkarmú (triungulinus) lárvák felkapaszkodnak a virágzó növényekre, és a porzószálak közt meglapulva várnak a gyanútlan beporzókra. Karmocskáikkal aztán az arra szálló méhek kitinszőreibe kapaszkodnak, és a fészekbe vitetik magukat. A célpontok jellemzően magányosan élő (szoliter) méhfajok, mint az Andrena, Anthophora és a Chalicodoma nemzetségek tagjai. A triungulinus lárva a méhsejtbe kerülve először az ott lévő tojást fogyasztja el, majd egy másik lárvastádiumot öltve a felhalmozott mézkészleteket kezdi felfalni. Mindeközben egy relatíve biztonságos helyen fejlődhet. Lenyűgöző és mindenekelőtt kényelmes életstratégia ez, a kleptoparazitizmus (más fajok zsákmányának, illetve önfenntartás vagy utódgondozás céljából felhalmozott készleteinek megdézsmálása) egy érdekes alesete.
Európai rokonainál egy fokkal erőszakosabb módon érnek célba egy amerikai nünükefaj, a Meloe franciscanus lárvái. Ez a faj az Egyesült Államok nyugati területeinek félsivatagos, sivatagos élőhelyein érzi magát igazán otthon. Egy-egy magasabb kórón a triungulinus lárvák egész kis tömege szokott összeverődni, egy magányos méhfaj nőstény egyedének sziluettjét és illatát utánozva. Ilyen magasszintű kooperációra euszociális rovarok körében bőséggel akad példa, de bogaraknál még csak hasonlót sem figyeltek meg eddig. Főleg egzotikus ez a viselkedési forma, mivel egy speciális mimikrivel is társul.
Egy szerelemtől vak, és párzásra kész hím méhet, ha sikerrel megtéveszt a Meloe franciscanus lárvák nyüzsgő kupacának látványa, és a lárvák által termelt feromonfelhő, amaz a párzás reményében túl közel merészkedik a lárvakupachoz, és ezzel kész is a baj. A lárvák megrohamozzák a méhet, és amennyien csak tudnak, belekapaszkodnak a bundájába. A testsúlyeloszlás hirtelen megváltozása miatt a lépre csalt méh gyakran drámai módon a földre zuhan, majd miután a lárvák kényelmesen elhelyezkednek rajta, és ő visszanyeri egyensúlyát tovább repül, hogy ezúttal egy valódi nőstényre akadjon. Miután méhünk sikeresen rátalált szíve választottjára, és beteljesült szerelmük, a nünükelárvák, mint valami nemi betegség, átragadnak a nőstényre. Innentől kezdve az ő bundáján laknak, és türelmesen várják, hogy a nőstény megépítse a fészket, lerakja petéit és megkezdje a méz felhalmozását. Ekkor aztán eljön a triungulinusok ideje! Belevetik magukat a mézbe, s innentől kezdve európai unokatestvéreikhez hasonló módon tobzódnak a jólétben.
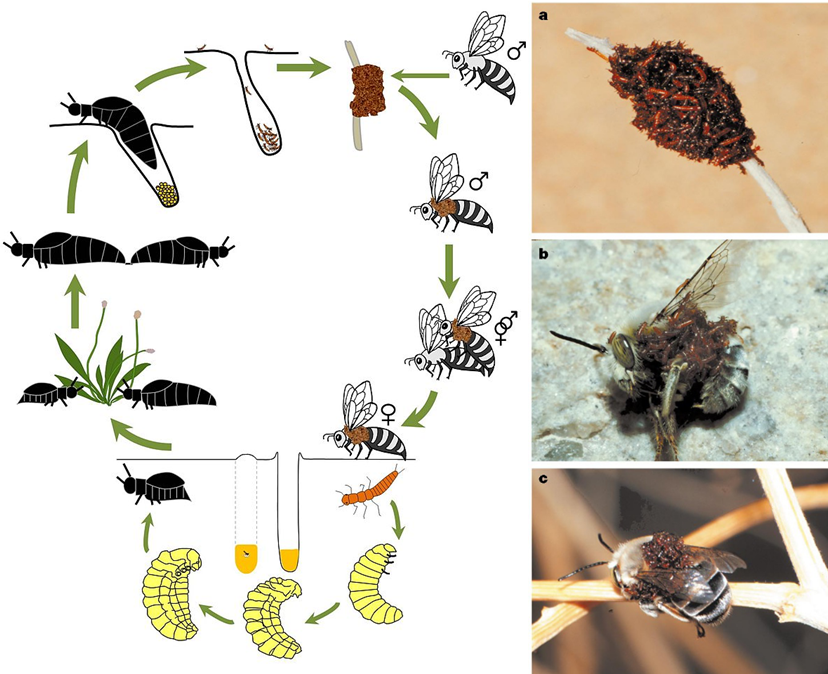
A Meloe franciscanus életciklusa (bal) (forrás: Wikipedia), triungulinus lárvák kupaca egy elszáradt növény szárán (a) és a megtévesztett méh testén (b; c) (forrás: Hafernik és Saul-Gershenz).
Belátható, hogy a M. franciscanus sikere azon áll, vagy bukik, hogy a lárvák magukhoz tudják-e vonzani a hím méhet. Minthogy ezt elsősorban az adott méhfaj nőstényeire jellemző feromonok termelésével érik el, a kleptoparazitizmus itt erősen gazdaspecifikus. A méhek ha szabadulni akarnak a nünükéktől, egyetlen evolúciós út járható számukra: változtatni kell egy kicsit a faj szexferomonjainak kémiai sajátosságain. Ez azonban könnyen reproduktív izolációhoz vezet az eredeti méhfajon belül, mivel az egyik populáció hímjei előbb utóbb nem lesznek képesek érzékelni egy másik populációba tartozó nőstény illatát. A populációk tehát előbb-utóbb faji szinten is elválnak egymástól.
Persze a parazitának is lépést kell tartania a „feromonkódok” újraírásával, ha nem akar gazda nélkül maradni. Erre szépen rímel az a megfigyelés, hogy a M. franciscanus Oregon part menti dűnéin élő populációja a Habropoda miserabilis nevű oregoni méhfajra specializálódott, míg ugyan ennek a nünükének a Mojave-sivatagban élő populációja előbbi méhfaj egy itt élő, közeli rokonfaján, a Habropoda pallida-n élősködik. A nünüke és a méhek közti kémiai verseny tetten érhető, ha (miként azt Leslie Saul-Gershenz és munkatársai tették) elemezzük az általuk termelt feromonkoktélok összetételét.
A hártyásszárnyúak szexferomonjai többnyire páratlan, 21-31 szénatomból álló, egyszeresen telítetlen szénhidrogének (a kettős kötés leggyakrabban a 9. szénatomnál található, de a különböző fajok feromonkoktéljaiban ettől eltérő pozícióba is kerülhet). A két Habropoda faj feromonjai között jellemző különbségeket találtak, és fajon belül természetesen a hímek és a nőstények ferromonprofilja is egészen másképp fest. Szépen tükrözi viszont a gazda-parazita kapcsolatot jellemző kémiai kódíró versenyt, hogy az oregoni M. franciscanus feromonjai között több olyan vegyület is található, ami a H. miserabilis nőstényére jellemző, míg a mojavei nünükék, noha biztosan ugyan abba a fajba tartoznak, mint az oregoniak (ezt anatómiai és genetikai elemzések is igazolják), feromonprofiljuk jelentősen eltérő. A mojavei populáció feromonkoktélja több olyan komponenst tartalmaz, ami a H. pallida-ra jellemző.

A H. miserabilis és a H. pallida hímjeinek és nőstényeinek valamint az oregoni és Mojave-sivatagi nünükelárvák feromon profilja. A vízszintes tengelyen a különböző szénatomszámú feromonok vannak feltüntetve, kék színnel jelölve azokat, amelyek legalább két csoportban ugyan úgy jelen vannak. a függőleges tengelyen relatív koncentráció látható (Leslie Saul-Gershenz és mtsai, 2017).
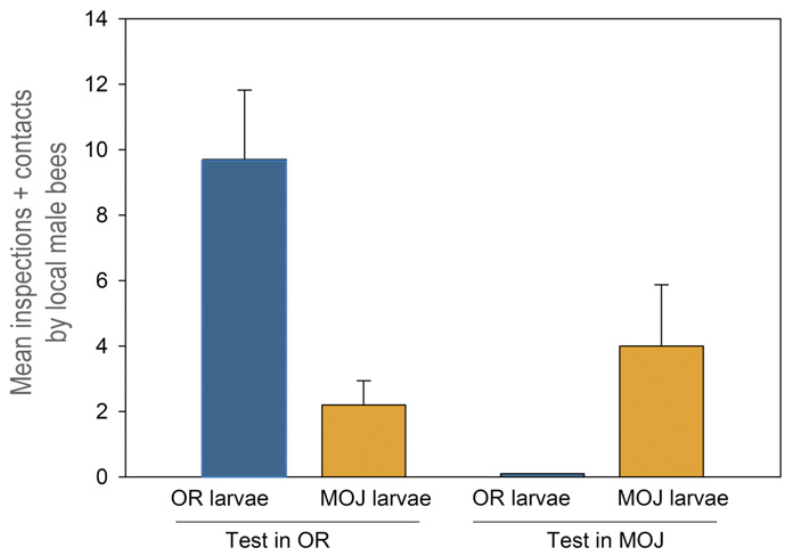
A feromonprofil vizsgálata mellett olyan kísérleteket is végetek a szerzők, amelyek a két nünükepopuláció eltérő gazdaspecificitását igazolták. Ezekből kiderült, hogy a mojavei nünükepopuláció valóban csak az ott élő Habropoda fajt vonzza magához, míg az oregonit pedig csak alig, vagy egyáltalán nem, és vice versa. Ezekről a kísérletekről olvasva az ember elgondolkozik rajta, hogy az eltérő gazdaspecificitás a két nünükepopuláció egyedei között (a jelentős földrajzi távolságokat nem számolva) mennyire járul hozzá a populációk szétválásához, vagy adott esetben magához a fajképződéshez…
A jelen poszt forrásául szolgáló tanulmányok nem a tudomány legfrissebb termékei, és még csak azt se hiszem, hogy világmegváltó jelentőséggel bírnának. Amiért mégis beszámoltam róluk, annak csak az az oka, hogy idehaza épp „nünüke szezon” van. Bár a hazai Meloe fajok életciklusa kevéssé bizarr mint az M. franciscanus-é, ha a nyájas olvasó a hétvégén netán kirándulásra adná a fejét, és teszem azt egy kék nünükével (M. violaceus) találkozna gondoljon majd ezekre az érdekes amcsi rokonaira is 😉
Szép időt, és kellemes túrázást!
(A borítóképen egy kék nünüke látható – a szerző fotója)
__________
John Hafernik és Leslie Saul-Gershenz (Nature, Brief Communcations, 2000): „Beetle larvae cooperate to mimic bees„
Leslie Saul-Gershenz és mtsai. (PNAS, 2017): „Deceptive signals and behaviors of a cleptoparasitic beetle show local adaptation to different hostbee species„