
 Ha Darwinról és hozzá közel álló madarakról beszélünk, akkor talán nem is feltétlenül a kötelező újggyakorlatként felemlegetett galapagosi pintyeket (aka. “Darwin pintyek”) kellene előhozzuk, hanem a galambokat.
Ha Darwinról és hozzá közel álló madarakról beszélünk, akkor talán nem is feltétlenül a kötelező újggyakorlatként felemlegetett galapagosi pintyeket (aka. “Darwin pintyek”) kellene előhozzuk, hanem a galambokat.
Nem sokkal “A fajok eredete” második kiadását követően (amiből szintén nem hiányoztak a galambok) publikálta Darwin „Az állatok és növények megváltozása a háziasítás állapotában” (The Variation of Animals and Plants under Domestication) c. könyvét, amiben két teljes fejezetet szentelt a különböző galambfajtáknak. Figyelembe véve, hogy Darwin maga is lelkes űzője volt kora állattenyésztési hobbijának, mindez talán nem is annyira meglepő, talán inkább az a releváns kérdés, hogy mennyire formálta a galambtenyésztés Darwin nézeteit az evolúcióról és hogyan használta kvázi modellélőlényként galambjait.
Mindenesetre, ha nem is pont így fogalmazott, Darwint saját keresztezései győzték meg arról, hogy a kedveltebb fajták kinézete tulajdonképpen olyan ritka természetes variációknak köszönhető, amelyek egyébként is jelen lehettek az ősi galambokban. És bár Darwin értelemszerűen még nem használta a domináns és recesszív fogalmakat, az általa leírt egyik kísérlet, amiben három fajtiszta vonal különböző hibridjeit keresztezte és végül az ősi formának tekintett szirti galambra emlékeztető utódok jöttek létre, tökéletesen példázza, hogy mi is történne, ha különböző gének recesszív alléljaira homozigóta (és a normálistól eltérő kinézetű) vonalakat egymással kereszteznénk.
Bár a galambászati kedv napjainkra kissé alább hagyott, azért még mindig kellő számban léteznek ezek a több száz év alatt létrehozott formák, hogy tanulmányozni tudjuk, mi is hozza létre a jellegzetes fenotípusos jegyeket, például a parókás galamb csuklyáját, vagy az angol trombitás tollas lábait. (Egyébként a mesterséges szelekció létrehozta formagazdagság, csak a kutyában és az aranyhalban tapasztalthoz hasonlítható, vagyis két olyan másik fajhoz, amelyeket szintén évszázadokon keresztül formált saját ízlése szerint az ember.)

Utóbbi azért is érdekes téma, mert a blog történetének egyik leggyakrabban elővett témájához vezet (ismét) vissza minket, ez pedig a végtagfejlődés. De menjünk sorban.
Az elmúlt években a galambok evolúciós modellélőlényként való felfedezése elsősorban a University of Utah-on dolgozó Michael Shapiro nevéhez köthető. Az ő csoportja volt például az, amelyik öt évvel ezelőtt egy borzasztó érdekes eLife cikkben a galambok (és tyúkok) tollas lábainak fejlődési eredetét próbálta feltárni.
Egy átlagos madárláb, ahogy azt mind tudjuk, pikkelyes, karmos – és szöges ellentétben az állatok mellső végtagjával, jellegzetesen tolltalan. De akkor mi történhetett mondjuk az angol trombitás esetében? Hogy megtaláljuk melyik gének lehetnek a fenotípus mögött, Shapiroék az egyik klasszikus, már Mendel által is alkalmazott “trükköt” vetették be: keresztezték a különböző kinézetű (fenotípusú) madarakat, létrehozhva egy hibrid generációt (F1), amelyeket egymás közt szaporítva immár egy olyan generációt kaphattak (F2), ahol a recesszív jelelgek is ismét meg tudtak nyilvánulni.
A modern genomika egyik előnye egyébként az is, hogy elég pontosan lehetett azt is látni, hogy melyek azok a DNS szakaszok, amelyek a két kiindulási fajta genomjának elvileg teljes összekeverése ellenére minden egyes tollaslábú F2 egyedben jelen lehetnek és megegyezenek a tollas lábakkal rendelkező kiindulási szülő esetében. Ezek rejtik jó eséllyel ugyanis azokat a mutációkat/szekvencia-variánsokat, amelyeknek köze lehet a furcsa lábak kialakulásához.
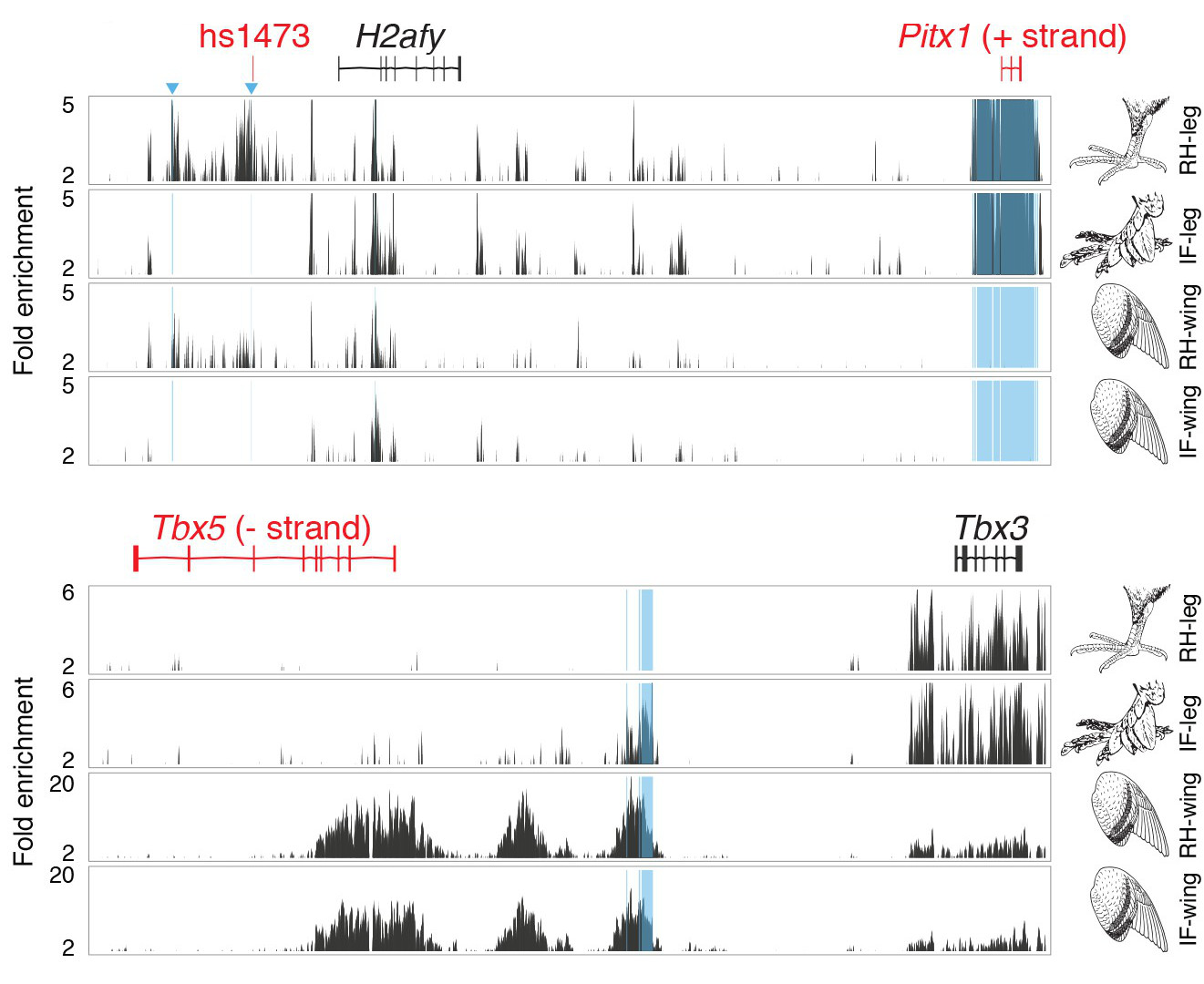
Ennek az eredményeként két fontos genomi pozíciót leltek, amelyek ráadásul a végtagfejlődésben eleve fontos gének közelében vannak: az egyik a Pitx1 gén mellett, a másik a Tbx5 és Tbx3 gének között helyezkedett el. Hogy ez miért felel meg kvázi a jackpot-megütésének egy ilyen típusú tanulmányban, ahhoz egyetlen sorát venném elő a már emlegetett végtagfejlődéses klasszikus posztnak:
A mezodermális szövet jellegét pedig három transzkripciós faktor határozza meg: a mellső végtagért a Tbx5, míg a hátulsóért a Tbx4 és a Pitx1 a felelős.
Azaz két olyan gén közelében láthatunk változásokat, amelyek terméke pont azt szabályozza, hogy a mellső és hátsó végtagok különbözőek legyenek. És a jelek szerint ez a mutációk hatására épp ez a különbség kezdett elmosódni a tollas lábak esetében.
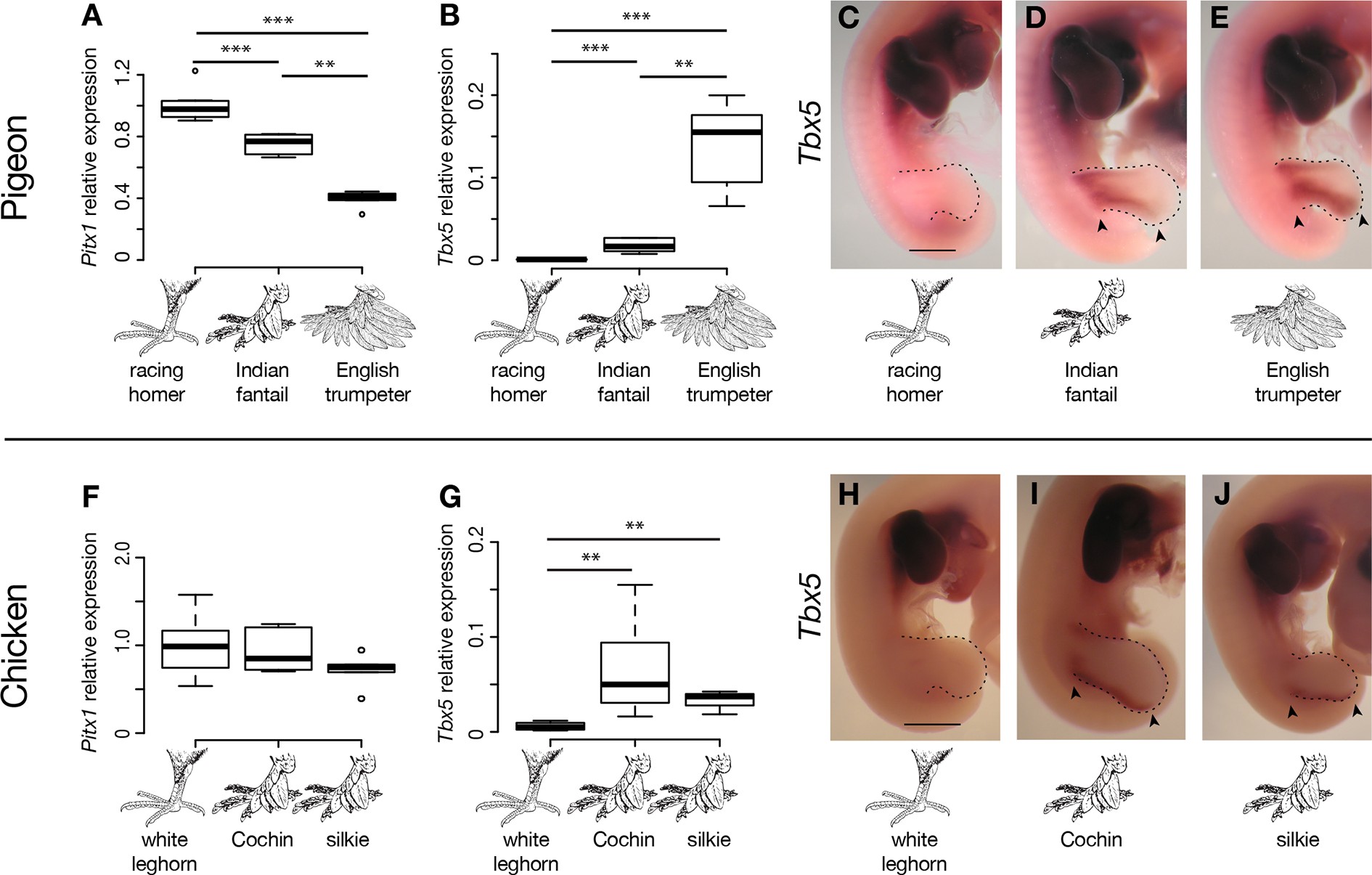
Ahogy a fenti ábra is jól mutatja: az angol trombitás esetében a hátsó végtag identitását biztosító Pitx1 expressziós szintje lecsökken, míg a normálisan csak a leendő szárnyakban expresszálódó Tbx5 hirtelen a hátsó végtagbimbóban is elkezd kifejeződni a fejlődés során. (S bár előbbi változás nem látható a gatyás tyúkokban, utóbbi annál is inkább.)
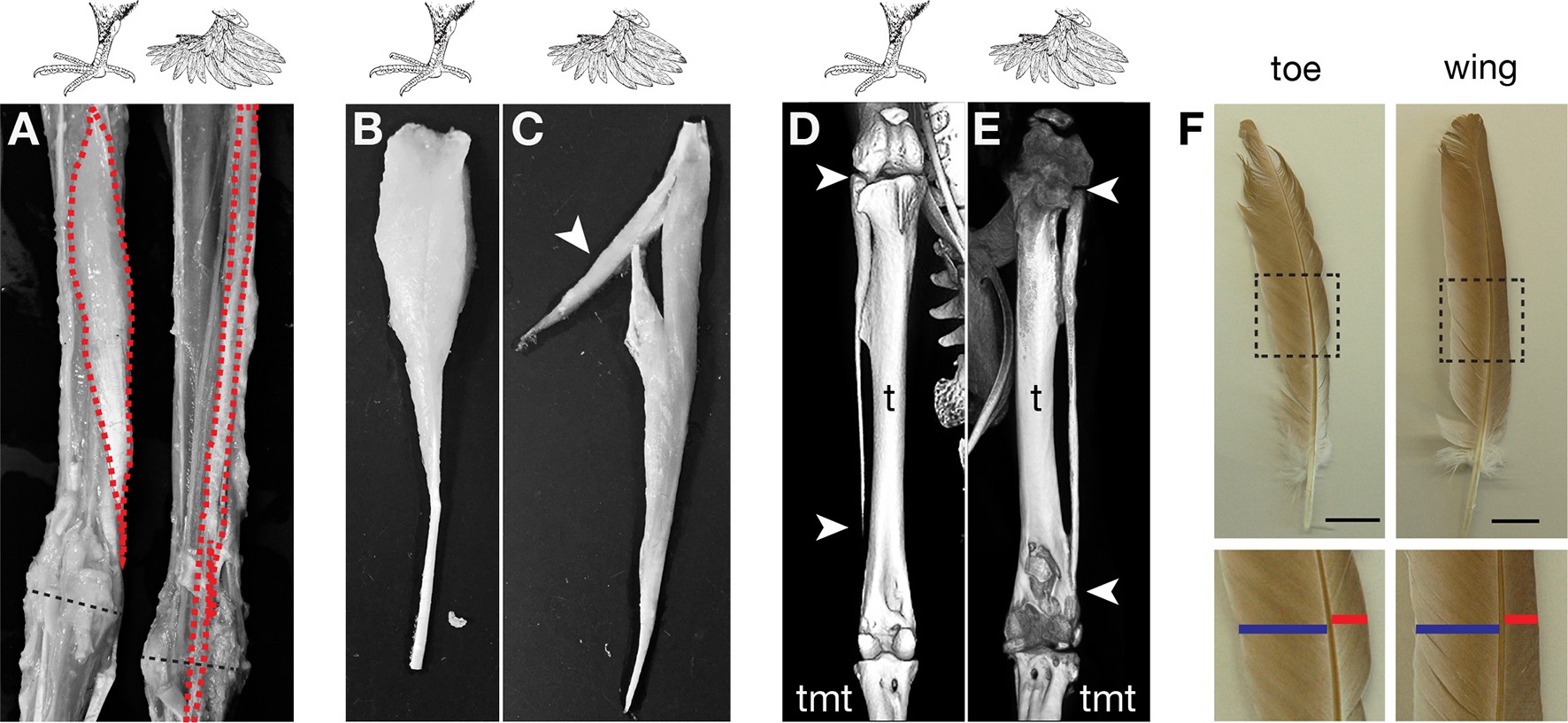
Ennek az ismeretnek a birtokában már talán kevésbé meglepő, hogy ha tüzetesebben megnézzük a tollas lábú galambok lábait, akkor egyrészt azt fedezhetjük fel, hogy ezek nem egyszerűen tollasok (és nem random tollakat, hanem a szárnyakra is jellemző fedőtollakat hordoznak), de alapvetően különböznek egy “normális” lábtól. Talán túlzás lenne azt állítani, hogy teljes egészében szárnyakká alakultak, de mindenképpen jól látható, hogy mind a csontok, mind az izmok alakja jellegzetesen különbözik a klasszikus madárlábtól, így egy kicsit “átmeneti” morfológiával rendelkező végtagot találunk itt és minden jel szerint ez az identitásvesztés, ami a tollazat megjelenését hozta magával.
Shapiroék csoportja azonban nem állt meg itt és pár további galamb-jelleg térképezésébe is belevágott, s ezekről kerültek fel preprintek a bioRxiv felületére.
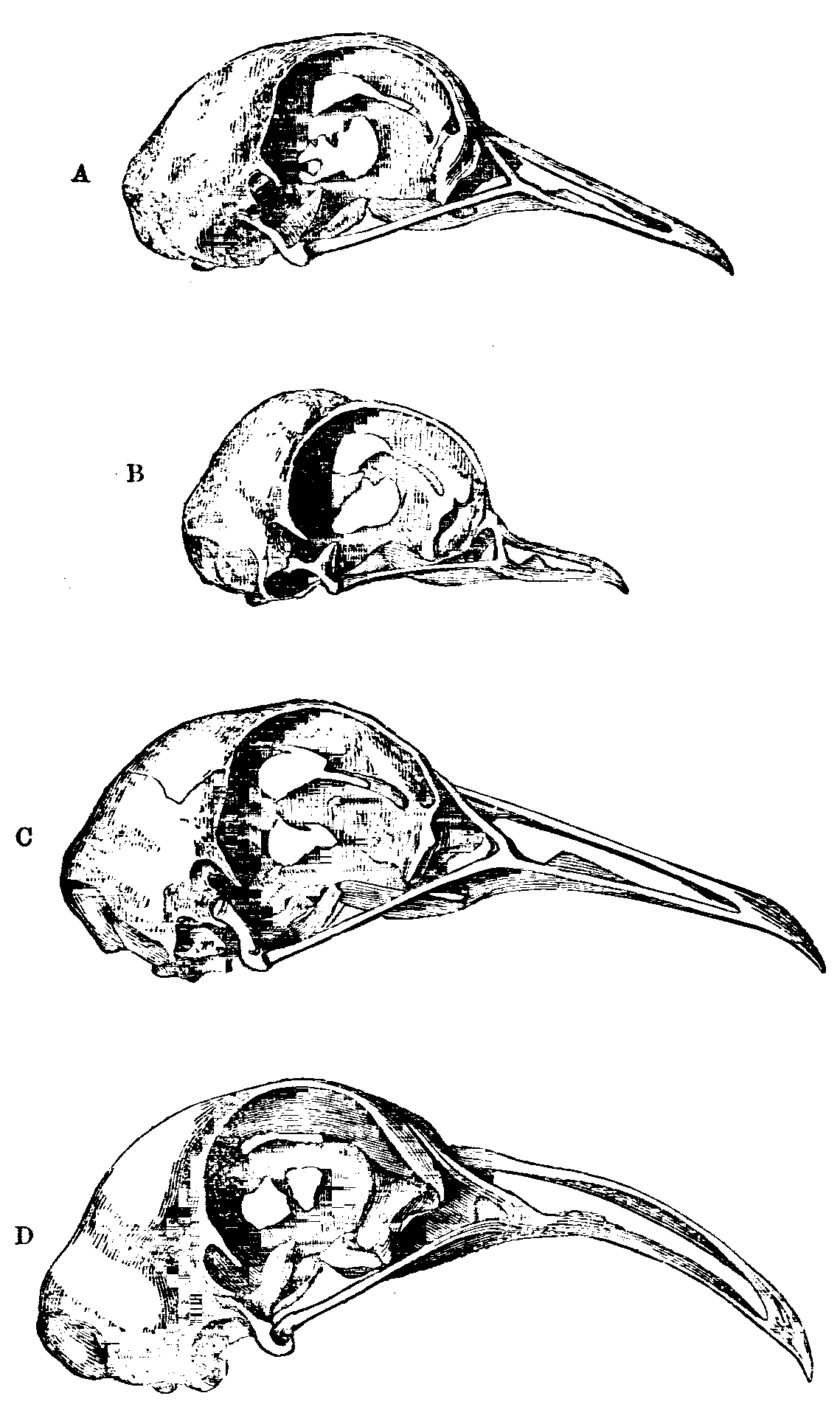
Az elsőben egy kicsit a galambkoponya alakjának furcsaságait próbálták megérteni. Ez egyébként szintén Darwin szívéhez közel álló téma, hiszen a „Az állatok és növények megváltozásában” maga is hosszan elemezgette ezt a dolgot.
A galambászat kiinduló fajának tekintett szirti galamb koponyájához képest, számod domesztikált fajtában, például az angol rövidcsőrű galamb esetében, látványosan megváltozott a koponya és csőr alakja és méretaránya. De talán sehol nem annyira látványos ez, mint az alsó állkapocs esetében, ahol akkora különbségek láthatók mondjuk a berber galambok, vagy az óriásra növő runtok esetében, hogy Darwin szavaival élve:
„Senki sem hitte volna el egykönnyen, hogy azok az állkapcsok, amelyek a fentebb meghatározott pontokban ennyire különböznek, egyazon fajhoz tartozhattak.”
Vagyis Shapiro csoportjának bőven volt miből válogatnia. Végül egy pomerániai begyes galambot és egy scandaroont kereszteztek, mert ezek csőrmorfológiája kellően különbözött, hogy egy kellően részletes jellemzéssel az F2 generációban már nagyon sok csőr-paraméter genetikai hátterét fel lehessen tárni.
A minél pontosabb jellemzéshez Shapiróék csoportja mikro-CT készülékek segítségével beszkennelte a madarak koponyáját és nagyon egzakt módon rengeteg paraméterét lemérték. S bár egyelőre, a tollas lábakhoz kapcsolódó tanulmányokkal ellentétben, ezek eredményeként nem tudunk konkrét géneket megnevezni, azért viszonylag pontosan meg lehet mondani, hogy melyik kromoszóma-régiókban rejtőznek a minket érdeklő genetikai variációk. (Ennek a metodológiáját valahogy úgy kell elképzelni, hogy néhány tucat F2 hibridnek megszekvenálják a genomját és lemérik a koponyáját a CT segítségével. Ezt követően pedig elkezdenek korrelációkat keresni bizonyos markerek jelenléte és bizonyos fizikai paraméterek között. Mivel a koponya- és csőrforma egyértelműen genetikailag kódolt, ilyeneknek kell lenniük. A kérdés ilyenkor inkább az, hogy elegendő-e ennyi egyed megszekvenálása, hogy ezeket mind megleljük – általánosságban, minél több gén befolyásol egy karaktert, annál több egyed szekvenciájára és méreteire van szükségünk az ilyen vizsgálatokban, hogy “észrevegyük”, hol bujkálnak a releváns mutációk.)
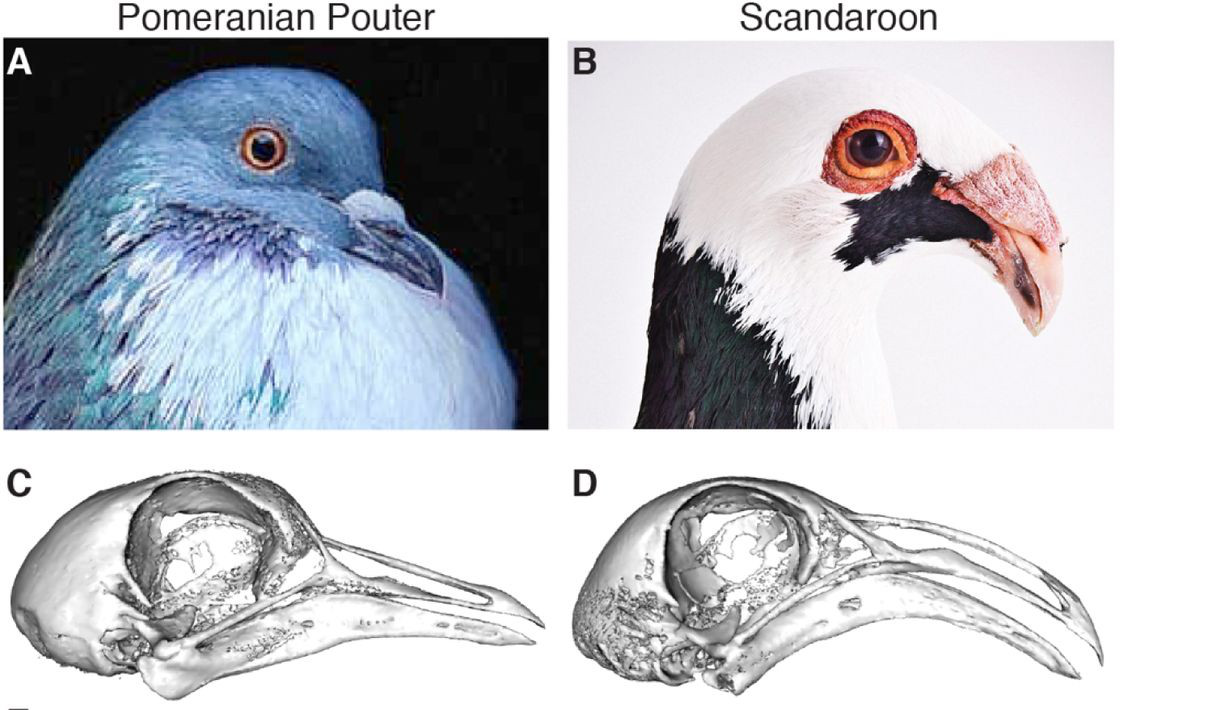
A koponya szélességének és hosszúságának legfontosabb paramétereit öt kromoszóma-régió befolyásolja, a koponya formáját pedig további hat-nyolc. Ez elég sok (például a galapagosi pintyeknél úgy néz ki, hogy csak két–három gén játszik szerepet a csőr evolúciójában), különösen mert a mostani adatok nem képesek egyetlen génre leszűkíteni a keresést, hanem kromozsóma szakaszokat tudnak csak megadni. És jelen pillanatban egy-egy ilyen régió akár több tucat gént is tartalmazhat, amelyek közül a legtöbb (még) nem lett sehogy a koponyaforma evolúciójához kötve más fajokban. Így aztán, bár részletes felsorolás adható, a szerzők is csak találgatni tudnak, hogy melyik génnek van valójában köze a csőr és koponya változásaihoz.
Ennél szerencsésebb a helyzet a szivárványhártya (írisz) esetében, ami normális esetben narancssárga színű a szirti galambnál, de vannak fajták, ahol gyöngyházszínű és megint máshol pedig színtelen, ami miatt a mögötte található retina fekete pigmentált epitéliuma adja majd a (fekete) színt.
A furcsa színek megértéséhez tudnunk kell, hogy a szemben három fajta pigment van jelen: a fekete melanin (a retinában), illetve a fehéres guanidin és a narancssárga pteridin a szivárványhártyában. Minden pigmens több lépéses bioszintézis útvonalak állítanak elő, melyeknél elég, ha az egyik elem megsérül és az adott pigmens (illetve az általa meghatározott szín) is eltűnik.

A galamboknál különböző keresztezések alapján úgy tűnik, hogy mind gyöngyházszínű (pearl), mind a fekete (bull) szemszínt valamilyen viszonylag egyszerű, monogénes jellegként lehet vizsgálni.
Ennek alapján narancssárga és gyöngyházszínű szemű szülők unokáit (F2) vizsgálva kiderült, hogy az okozati mutáció jó eséllyel egy SLC2A11B nevű génben keresendő, ugyanis a fehéres szemű egyedekben ez a gén egy olyan mutációt hordoz, ami miatt terméke rövidebb és működésképtelen lesz. Viszonylag keveset tudunk erről a konkrét génről, eltekintve, hogy a sejtmembránban levő szállító funkciót betöltő fehérjét kódol és az ilyen típusú géneknek egyébként is szerepe van mindenféle állat pigmentációjában (igen, a miénkben is), a medaka nevű ázsiai halfajban konkrétan a pteridin-szintézishez is köze lehet.
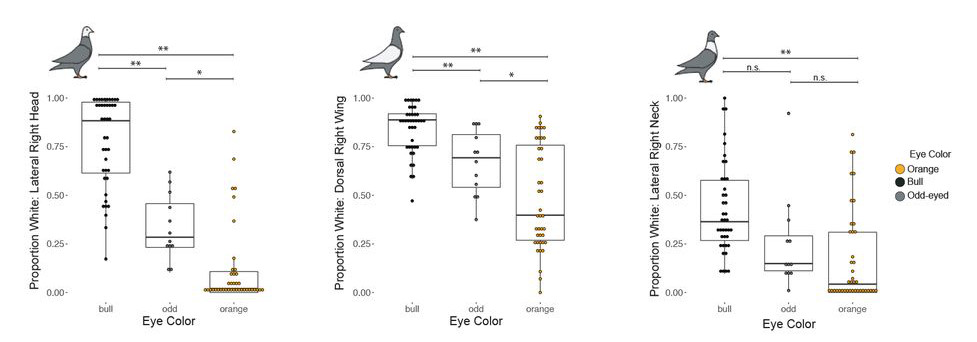
De a SLC2A11B csak fela a történetnek, a pigmentáció teljes hiányát a bull-szemű egyedekben más génnek a számlájára kell/lehet írni. Ez pedig, legavlószínűbb, hogy az EDNRB2 nevű, endothelin-receptort kódoló gén lehet. Tény, hogy itt nem azonosítottak olyan mutációt a kutatók a fekete szemű egyedekben, ami a gén működésképtelenségére utalna, a releváns genetikai régióban ez az egyetlen olyan gén, aminek köze van a pigmentációhoz, ráadásul számos más háziszárnyasban bizonyították be, hogy EDNRB2 mutációi depigmentációt eredményeznek.
Ami azért is érdekes, mert úgy tűnik, hogy a – feltehetőleg – szabályozó mutáció hatása nem szorítkozik egyedül a szemre, és ugyanazok az egyedek, amelyek bull-típusú szemet hordoznak, testük számos egyéb területén depigmentáltak: a fejükön szinte mindig, de nagyon gyakran a szárnyukon és a nyakukon is.
Szóval itt állunk most, de még biztos, hogy ahogy a kutyák is szinte kimeríthetetlen forrásai a genetikai kutatásoknak, a galambokról is fogunk jópárszor hallani.
Domyan ET, Kronenberg Z, Infante CR, Vickrey AI, Stringham SA, et al. (2016) Molecular shifts in limb identity underlie development of feathered feet in two domestic avian species. eLife 5: e12115. doi: 10.7554/eLife.12115.
Boer EF, Maclary ET, Shapiro MD (2021) Complex genetic architecture of three-dimensional craniofacial shape variation in domestic pigeons. bioRxiv doi: 10.1101/2021.03.15.435516.
Maclary ET, Phillips B, Wauer R, Boer EF, Bruders R, et al. (2021) Two Genomic Loci Control Three Eye Colors in the Domestic Pigeon (Columba livia). bioRxiv doi: 10.1101/2021.03.11.434326.