
 A négylábú gerincesek fejlődésének ezernyi érdekfeszítő része közül az egyik legérdekesebb és ennek megfelelően legtöbbet tanulmányozott, a végtagok kialakulása. Az állatok ezen csoportjának nevét (Tetrapoda) is adó függelékek azért különösen izgalmasak egy fejlődésbiológus szemszögéből, mert rajtuk keresztül az alapoktól megérthető, hogy miként jöhet létre egy komplex szerv a maga térbeli elrendeződésével és milyen jelátviteli mechanizmusok kellenek az egyes komponenseinek kialakításához. (Gondoljunk csak a saját kezünkre, hányfajta csont, ér, ín, ideg és izom van benne, illetve, hogy jól láthatóan három térbeli tengely mentén szerveződik.) Ráadásul a végtagok a fejlődő állat szempontjából nem is létfontosságúak, így kíséreletek során viszonylag szabadon manipulálhatjuk őket, ami nem elhanyagolható a kutató önös szempontjából sem.
A négylábú gerincesek fejlődésének ezernyi érdekfeszítő része közül az egyik legérdekesebb és ennek megfelelően legtöbbet tanulmányozott, a végtagok kialakulása. Az állatok ezen csoportjának nevét (Tetrapoda) is adó függelékek azért különösen izgalmasak egy fejlődésbiológus szemszögéből, mert rajtuk keresztül az alapoktól megérthető, hogy miként jöhet létre egy komplex szerv a maga térbeli elrendeződésével és milyen jelátviteli mechanizmusok kellenek az egyes komponenseinek kialakításához. (Gondoljunk csak a saját kezünkre, hányfajta csont, ér, ín, ideg és izom van benne, illetve, hogy jól láthatóan három térbeli tengely mentén szerveződik.) Ráadásul a végtagok a fejlődő állat szempontjából nem is létfontosságúak, így kíséreletek során viszonylag szabadon manipulálhatjuk őket, ami nem elhanyagolható a kutató önös szempontjából sem.
A történet egy sima testfallal kezdődik, amelyen egyelőre nyoma sincs annak, hogy hamarosan egy hosszú, hengeres valami lóg majd ki belőle. De molekulárisan a testfal egyes pontjai, sejtcsoportjai már elköteleződtek hogy végtagokká váljanak (ha pl. egy szalamadra embrióban a szóbanforgó területet teljesen nem adkevát helyekre – mondjuk fej – ültetve, ott végtagokat fogunk kapni).
Az első izgalmas kérdés, hogy honnan „tudja” a fejlődő embrió testfala, hogy adott magasságban „illik” egy pár kezet vagy lábat kialakítania? A válasz a mai tudásunk szerint a test hosszanti felosztásáért felelős Hox-kódban rejlik (egyébként a Hox gének történetünk visszatérő szereplői lesznek, itt frissítheti fel a tudását mindenki róluk). Mint arról a blog legelső postjában már írtam, a mellső végtag helye a nyaki és mellkacsi csigolyák találkozásánal van, ami egybeesik a hoxc6 gén kifejeződési tartományának elejével. (A hátsó végtag helyezkedése még kevésbé ismert.)
 A tesfal ily módon megjelölt pontjaiban a külső csíralemez egy különleges tarajszerű képződmény hoz létre, az apikális ektodermális redőt (AER – apical ectodrmal ridge), amely majdnem a fejlődési folyamat végéig a végtag távolabbi, ún. disztális részét borítja majd. Az AER sejtjei az FGF (fibroblast growth factor) osztályba tartozó növekedési faktorokat termelnek, amelyeknek fontos szerepük van abban, hogy az alattuk levő sejteket osztódásra késztessék. Ennek következtében előbb egy kidudorodás, majd egy egyre jobban látható ún. végtagbimbó kezd kialakulni. A bimbó belsejében lezajló folyamatok hatására egyes sejtekből porcsejtek lesznek, amelyek összetömörödve létrehozzák a későbbi csontok mintájául szolgáló porcos szöveteket (ebben elévülhetetlen érdemei vannak az egyes bmp géneknek – lásd még miért is lesz a denevér szárnya olyan amilyen) illetve a köztük levő ízületeket. (A folyamat a végtagbimbó testfalhoz közelebbi, azaz proximális részében zajlik le először és a végtag fejlődése során fokozatosan terjed tovább.) Az izmok a testfal leendő izomkötegeit kialakító szomitákból később indevándorló mezodermális sejtekből jönnek létre.
A tesfal ily módon megjelölt pontjaiban a külső csíralemez egy különleges tarajszerű képződmény hoz létre, az apikális ektodermális redőt (AER – apical ectodrmal ridge), amely majdnem a fejlődési folyamat végéig a végtag távolabbi, ún. disztális részét borítja majd. Az AER sejtjei az FGF (fibroblast growth factor) osztályba tartozó növekedési faktorokat termelnek, amelyeknek fontos szerepük van abban, hogy az alattuk levő sejteket osztódásra késztessék. Ennek következtében előbb egy kidudorodás, majd egy egyre jobban látható ún. végtagbimbó kezd kialakulni. A bimbó belsejében lezajló folyamatok hatására egyes sejtekből porcsejtek lesznek, amelyek összetömörödve létrehozzák a későbbi csontok mintájául szolgáló porcos szöveteket (ebben elévülhetetlen érdemei vannak az egyes bmp géneknek – lásd még miért is lesz a denevér szárnya olyan amilyen) illetve a köztük levő ízületeket. (A folyamat a végtagbimbó testfalhoz közelebbi, azaz proximális részében zajlik le először és a végtag fejlődése során fokozatosan terjed tovább.) Az izmok a testfal leendő izomkötegeit kialakító szomitákból később indevándorló mezodermális sejtekből jönnek létre.
Kicsit időben előre futva, ha megnézünk egy kifejlett végtagot (az egyszerűség kedvéért maradjunk ismét a kezünknél), láthatjuk amint a válltól távolodva fokozatosan különböző csontok (és izmok) helyezkednek el. Ez az ún. proximo-disztális (ProDis) végtagtengely, amelyen a felkar a stylopod, az alkar a zygopod, a tenyér és az ujjak pedig az autopod régiót alkotják. Jelenleg az egyes régiók létrejöttére vonatkozóan két nézet létezik.
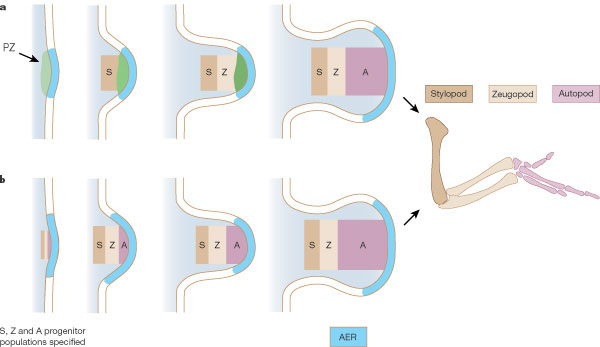
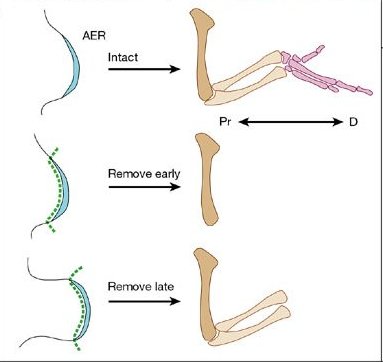 Az egyik a Lewis Wolpert nevéhez fűződő, egy haladási zóna (PZ – progress zone) létét posztuláló gondolat, amely értelmében az, hogy egy bizonyos sejt mennyi időt tölt ebben, az AER alatt fekvő zónában meghatározza, hogy milyen régió identitását veszi fel (a legtovább az autopod sejtjei vesztegelnek a PZ-ban). Elég sok adat látszik a PZ létét alátámasztani: pl. az AER-t minnél hamarabb távolítjuk el, annál több disztális régió hiányzik majd. A másik elmélet, a korai specifikáció elmélete (early specification model) elmélete, amely hívei szerint az egyes régiók már a fejlődés legelejétől léteznek. Ezt látszik alátámasztani az, hogy a korai végtagbimbó különböző mélységeiben megjelölt sejtek jól megjósolhatóan a mélységgel korelállóan kötnek majd ki adott régióban. Azonban ez a megfigyelés önmagában nem mond ellent feltétlenül az első, klasszikus nézetnek, hiszen az is lehet, hogy a korai végtagbimbó mélysége azt is jelenti egyben, hogy egy-egy sejt milyen hamar léphet majd ki a PZ-ból. (Talán nem kell ragozni, hogy egyelőre szerintem Wolpert elmélete tűnik jobbnak ;-)).
Az egyik a Lewis Wolpert nevéhez fűződő, egy haladási zóna (PZ – progress zone) létét posztuláló gondolat, amely értelmében az, hogy egy bizonyos sejt mennyi időt tölt ebben, az AER alatt fekvő zónában meghatározza, hogy milyen régió identitását veszi fel (a legtovább az autopod sejtjei vesztegelnek a PZ-ban). Elég sok adat látszik a PZ létét alátámasztani: pl. az AER-t minnél hamarabb távolítjuk el, annál több disztális régió hiányzik majd. A másik elmélet, a korai specifikáció elmélete (early specification model) elmélete, amely hívei szerint az egyes régiók már a fejlődés legelejétől léteznek. Ezt látszik alátámasztani az, hogy a korai végtagbimbó különböző mélységeiben megjelölt sejtek jól megjósolhatóan a mélységgel korelállóan kötnek majd ki adott régióban. Azonban ez a megfigyelés önmagában nem mond ellent feltétlenül az első, klasszikus nézetnek, hiszen az is lehet, hogy a korai végtagbimbó mélysége azt is jelenti egyben, hogy egy-egy sejt milyen hamar léphet majd ki a PZ-ból. (Talán nem kell ragozni, hogy egyelőre szerintem Wolpert elmélete tűnik jobbnak ;-)).
A végtagrégiók identitását végsősoron az AER-ből, azaz disztálisan eredő FGF-, illetve a testfal felől, azaz proximálisan eredő retinálsav (RA) jelgradiens összjátéka határozza meg – utóbbi a Meis1 és Meis2 géneken keresztül hatva. (Persze más folyamatok is szerepet játszanak, de ma az említett kettő tűnik a legfontosabb szereplőnek.) A két, ellentétes forrású, s így kvázi komplementer erősségű gradiens régi ismerőseinken, a Hox géneken keresztül fejti ki hatását.
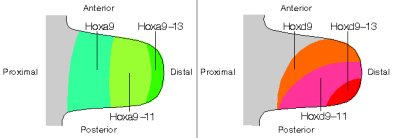 Mint az a mellékelt ábrán látható, két Hox-kluszter, az -a és a -d egyaránt kollineárisan fejeződik ki a végtagbimbóban és ez a kollinearitás valóban befolyásolja az egyes régiók identitását: a Hoxa9 és Hoxd9 (avagy rövidebben Hox9 paralógok) a váll ill. lapocka (scapula), a Hox10 paralógok a felkarcsont (humerus), a Hox11 az alkarcsontok (ulna és radius), a Hox12 a kéztőcsontok (carpals), a Hox13 pedig a kézközépcsontok (metacarpals) és ujjak mibenlétét határozzák meg. Például azokban az egerekben, amelyekben mind a Hoxa11, mind a Hoxd11 génre mutánsak, hiányoznak az alkarcsontok – lásd az alábbi ábra jobb oldalán -, a Hoxa13 és Hoxd13 együttes működésképtelensége pedig az ujjak elvesztését okozza.
Mint az a mellékelt ábrán látható, két Hox-kluszter, az -a és a -d egyaránt kollineárisan fejeződik ki a végtagbimbóban és ez a kollinearitás valóban befolyásolja az egyes régiók identitását: a Hoxa9 és Hoxd9 (avagy rövidebben Hox9 paralógok) a váll ill. lapocka (scapula), a Hox10 paralógok a felkarcsont (humerus), a Hox11 az alkarcsontok (ulna és radius), a Hox12 a kéztőcsontok (carpals), a Hox13 pedig a kézközépcsontok (metacarpals) és ujjak mibenlétét határozzák meg. Például azokban az egerekben, amelyekben mind a Hoxa11, mind a Hoxd11 génre mutánsak, hiányoznak az alkarcsontok – lásd az alábbi ábra jobb oldalán -, a Hoxa13 és Hoxd13 együttes működésképtelensége pedig az ujjak elvesztését okozza.

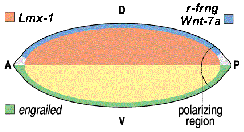 Ennek a tengelynek a kialakulása összefügg a test DV tengelyének alakulásával és az utóbbit kialakító folyamatok vezetnek azokhoz a molekuláris asszimetriákhoz, amelyek a kezünkben kialakítják az alaktani különbségeket. A lényegre térve, a dorsális (azaz kézeji) oldal ektodermális, külső rétegében fejeződik ki a wnt-7a és a radical fringe (rfng) gén. Előbbi az alatta levő sejtekben kiváltja az lmx-1 gén kifejeződését (amely dorsális mesodermális jelleget ad ezeknek a sejteknek) , míg utóbbi a ventrális (azaz tenyér felőli) oldalon komplementer módon kifejeződő engrailed (en) génnel együtt, a két expressziós terület határán az AER-t alakítja ki. A wnt-7a és en között kölcsönös antagonizmus áll fenn, egymás expresszióját szűkítik a már említett területekre. Emellett, a wnt7a (és egyúttal lmx-1) fontosságát jól tükrözi az is, hogy azokvan a wnt-7a -/- kisegerekben, amelyekben a gén nem működik kézfej helyett is tenyér jellegű struktúra jön létre.
Ennek a tengelynek a kialakulása összefügg a test DV tengelyének alakulásával és az utóbbit kialakító folyamatok vezetnek azokhoz a molekuláris asszimetriákhoz, amelyek a kezünkben kialakítják az alaktani különbségeket. A lényegre térve, a dorsális (azaz kézeji) oldal ektodermális, külső rétegében fejeződik ki a wnt-7a és a radical fringe (rfng) gén. Előbbi az alatta levő sejtekben kiváltja az lmx-1 gén kifejeződését (amely dorsális mesodermális jelleget ad ezeknek a sejteknek) , míg utóbbi a ventrális (azaz tenyér felőli) oldalon komplementer módon kifejeződő engrailed (en) génnel együtt, a két expressziós terület határán az AER-t alakítja ki. A wnt-7a és en között kölcsönös antagonizmus áll fenn, egymás expresszióját szűkítik a már említett területekre. Emellett, a wnt7a (és egyúttal lmx-1) fontosságát jól tükrözi az is, hogy azokvan a wnt-7a -/- kisegerekben, amelyekben a gén nem működik kézfej helyett is tenyér jellegű struktúra jön létre.
Tovább lépve elérkezünk a harmadik, azaz anterior-posterior (AP) végtagtengelyhez, amely leegyszerüsítve a hüvelyk- és kisujjunk között fut (persze nem csak ott, de ez a leglátványosabb).
Már a korai klasszikus embriológiai kísérletek során nyilvánvalóvá lett, hogy egy, a végtagbimbó hátsó részén levő sejtcsoport felelős a kéz polaritásának kialakulásáért. Ha egy csirkeembrióból kivágjuk a szóbanforgó kis szövetdarabot és egy másik emrbió végtagbimbójának elejébe ültetjük, a létrejövő „kézben” az elülső rész a hátsó rész tükörképe lesz (mintha a hüvelyk- és mutatóujj helyett is gyűrűs- és kisujj keletkezne). Ezen tulajdonsága miatt a végtagbimbó szóbanforgó részét a polarizáló aktivitás zónájának (zone of polarizing activity – ZPA) nevezik. Hosszú évekig homályban maradt, hogy melyik gén felelős ezért az aktivitásért, de ma már tudjuk, hogy a főszereplő a sonic-hedgehog (shh) nevű gén.
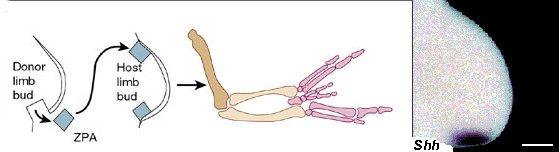
Jogosan vetődhet fel a kérdés, hogy vajon miért expresszálódik a shh pont ezen a területen? A válasz gyakorlatilag az, hogy mind a ProDis-, mind a DV mintázatot létrehozó molekulák összjátékának eredményeként. Az AER által termelt FGF molekulák, a dorsális ektodermában jelen levő Wnt-7a, a test RA gradiense és a végtagbimbóban kifejeződő Hox-d kluszter (amelynek, mint azt egy fenti ábrán látni lehet, a kollineáris expressziója szintén el van egy kicsit torzulva az AP tengely mentén) egyaránt közreműködik ebben. Ha valamelyik komponenst elvesszük, az óhatatlanul is a shh expressziójának hiányához vezet.
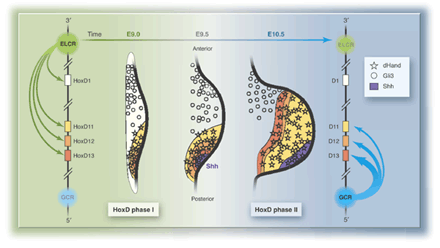 Külön érdekesség, hogy a shh expresszió egyszerre függ a Hox-d gének kifejeződésétől, és ugyanakkor maga is szabályozója a Hox-d géneknek. Pontosabban, miután a fejlődés kezdeti szakaszaiban a Hox-d gének hozzájárulnak a shh expresszió elindításához a Hox-d13 kifejeződési területén, a következő fázisban a shh maga aktiválja a Hox-d kluszter egy másik promóterét. Ennek következtében a Hox-d gének expressziós kollinearitása hirtelen a feje tetejére áll: shh kifejeződési helyén az előzőleg viszonylag távolabb expresszálódó Hox-d11 aktiválódik és a Hox-d13 kerül a legtávolabb. A fejük tetejére állított Hox-d géneknek ebben a fázisban az ujj-identitás kialakításában kell fontos szerepet játszaniuk.
Külön érdekesség, hogy a shh expresszió egyszerre függ a Hox-d gének kifejeződésétől, és ugyanakkor maga is szabályozója a Hox-d géneknek. Pontosabban, miután a fejlődés kezdeti szakaszaiban a Hox-d gének hozzájárulnak a shh expresszió elindításához a Hox-d13 kifejeződési területén, a következő fázisban a shh maga aktiválja a Hox-d kluszter egy másik promóterét. Ennek következtében a Hox-d gének expressziós kollinearitása hirtelen a feje tetejére áll: shh kifejeződési helyén az előzőleg viszonylag távolabb expresszálódó Hox-d11 aktiválódik és a Hox-d13 kerül a legtávolabb. A fejük tetejére állított Hox-d géneknek ebben a fázisban az ujj-identitás kialakításában kell fontos szerepet játszaniuk.
Hogy teljes legyen a kép, fontos hangsúlyozni, hogy a ZPA elsősorban az alkar és a kéz AP tengelyének definiálásában fontos, a felkar, ill. a váll ugyanezen tengelyét a test AP tengelyének koordinálását is végző RA gradiens hozza létre.
Eddig tehát végigvettük, hogy mely folyamatok hozzák létre a kezünk mintázatát a három térbeli tengely mentén. Még annyit érdemes megjegyezni, hogy az ezen a ponton az ujjak között levő szöveteknek (amelyek a legtöbb szárazföldi négylábúban később elpusztulnak, míg a vízben élőkben az úszóhártyát hozzák majd létre) abban van pontos szerepe, hogy az ujjpercek számát szabályozzák.
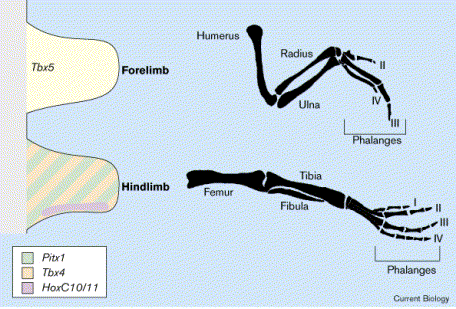 Látszólag a sztori végére értünk, van azonban még egy fontos kérdés. Minden hasonlóságuk ellenére, azért az első pár és a hátsó pár végtagunk különbözik egymástól. Miért van ez?
Látszólag a sztori végére értünk, van azonban még egy fontos kérdés. Minden hasonlóságuk ellenére, azért az első pár és a hátsó pár végtagunk különbözik egymástól. Miért van ez?
A korai embriológiai átültetéses kísérletek már rávilágítottak arra, hogy a végtagbimbó mezodermája felelős a végtag karakterének kialakításáért. Pl., ha a hátsó végtagbimbó mesodermáját egy mellső végtagbimbó mezodermája helyére ültetjük, akkor kéz helyett láb alakul ki. A mezodermális szövet jellegét pedig három transzkripciós faktor határozza meg: a mellső végtagért a Tbx5, míg a hátulsóért a Tbx4 és a Pitx1 a felelős.
Szóval ennyi lenne dióhéjban a négylábú gerincesek végtagkialakulásának fejlődésbiológiája. Kicsit hosszú lett, de mivel a közeljövőben számos, ehhez kapcsolódó dolgról szeretnék írni, jobbnak tűnt egy ilyen összefoglalót megereszteni. Ha vannak homályos pontok (ilyen zanzásított összefoglalók esetén ez valszleg elkerülhetetlen) azt a kommenteknél kéretik jelezni.
Végezetül még annyi érdekesség, hogy egy emberi magzat esetében a fent vázolt folyamatok röpke harmic nap alatt játszódnak le (a 26. napon kezdődnek) és a két hónapos magzat már egy teljesen megformált, bár mindössze 1 cm hosszúságú karral és lábbal rendelkezik. Innentől kezdve a terep a már „csak” a méretbeli növekedésé.
Wolpert, L. (ed.) Principles of Development. (ISBN 0198792913)
Gilbert, S. (ed.) Developmental Biology – Chapter 16. (ISBN 0878932585)
Mariani FV, Martin GR. (2006) Deciphering skeletal patterning: clues from the limb. Nature 423: 319-25.
Deschamps, J (2004) Hox Genes in the Limb: A Play in Two Acts. Science 304: 1610-1611.
Zákány, J, Kmita, M, Duboule, D (2004) A Dual Role for Hox Genes in Limb Anterior-Posterior Asymmetry Science 304: 1669-1672.
Tickle C. (2006) Making digit patterns in the vertebrate limb. Nat Rev Mol Cell Biol 7(1): 45-53.
Tickle C. (2003) Patterning systems–from one end of the limb to the other. Dev Cell 4(4): 449-458.
Graham, A. and McGonnell. I. (1999) Limb development: Farewell to arms. Current Biology 9(10):R368-370.
Koussoulakos S. (2004) Vertebrate limb development: from Harrison’s limb disk transplantations to targeted disruption of Hox genes. Anat Embryol (Berl). 209(2): 93-105.
University of Utah – DevBio course
Bár az államvizsga előtt találtam volna ezt meg…
roppant erdekes 🙂
pedig en informatikus vagyok 🙂
Az ujjak hosszarányának kialakulásáról is jó lenne majd egy cikk. 🙂
A kézfej helyett (ami az egész struktúrát jelenti)a megfelelő környezetben javaslom a kézhát szót a kézfej dorsalis oldalának megnevezésére.
Jó leírás, gratulálok!
„A következő az ún. dorso-ventrális (DV) végtagtengely, amely kvázi a kézfej-tenyér tengelynek felel mellső végtagunk viszonyában.”
Szerencsésebb lenne dorso-voláris tengelyről beszélni, (esetleg kézhát-tenyér-tengelyről).
Kéz”fej” (és láb”fej”) szakmailag nem jó kifejezés, az csak kéz és láb. (Van felső végtag, felkar alkar, kéz…)
Mint ahogy gerinc”oszlop” sincs, csak gerinc (mert mi olyan szerencsések vagyunk, hogy nem azt mondjuk – mint más nyelvek -, hogy csigolyaoszlop….).
Ha Vitrainak sikerült a „null” szót nullára alakítania (a fiatalok erre már nem is emlékezhetnek), bízom benne, hogy ezek is letűnnek egyszer a használatból.