
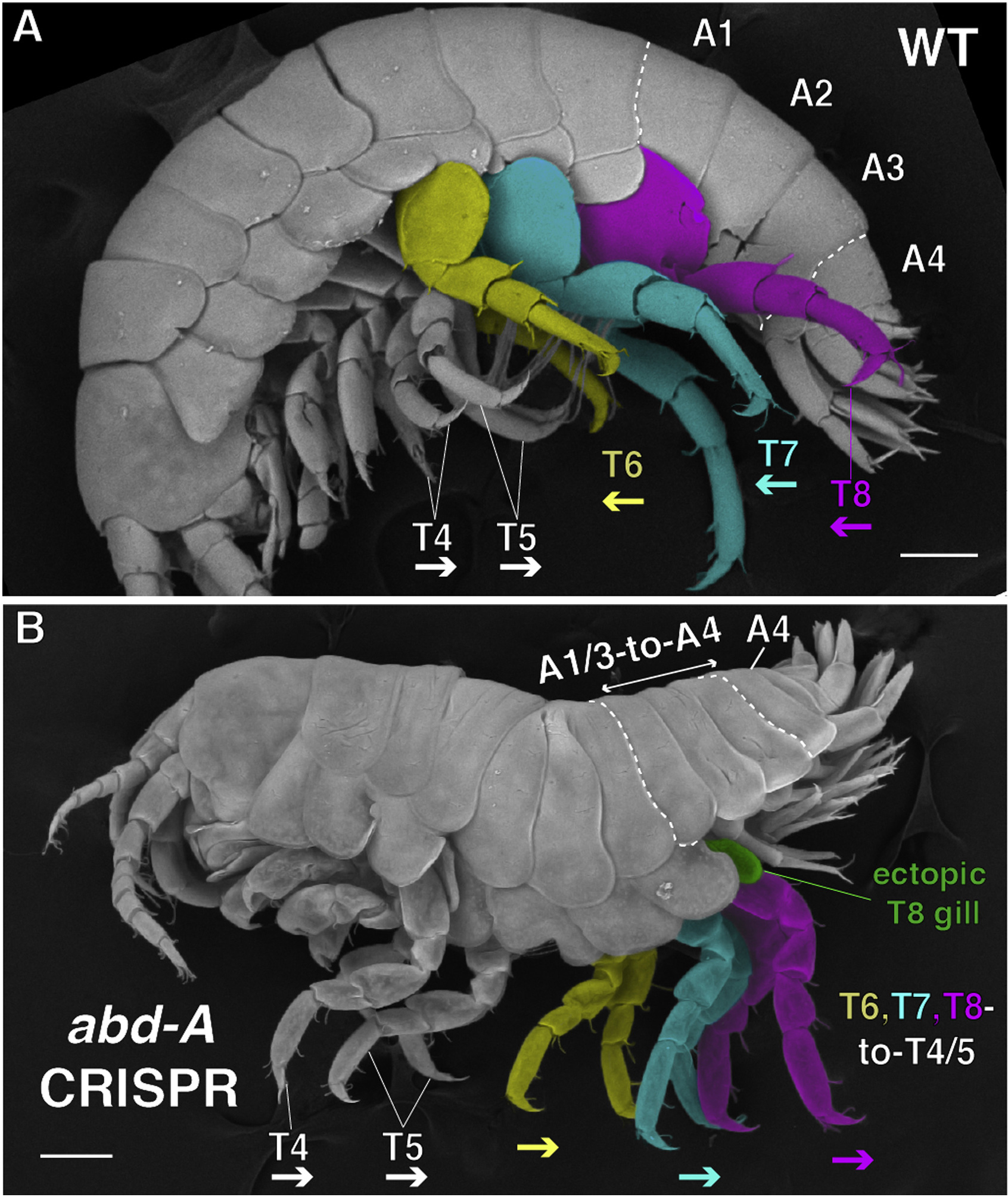
A bogarak rendje (Coleoptera) az egyik legnagyobb az élővilágban és a bennük fellelhető fantasztikus diverzitás eredete már évtizedek óta lenyűgözi a kutatókat.

A bogarak rendje (Coleoptera) az egyik legnagyobb az élővilágban és a bennük fellelhető fantasztikus diverzitás eredete már évtizedek óta lenyűgözi a kutatókat.
Az érdekes cikkek gyakran párban jönnek. Alig pár hete írtam a Cell-ben megjelent tanulmányról, amelyben a kígyók hátsó végtagjának eltünésében fontos szabályozó szakaszt mutattuk be és máris egy remek folytatást, pontosabban kiegészítést lehet hozzá tenni.
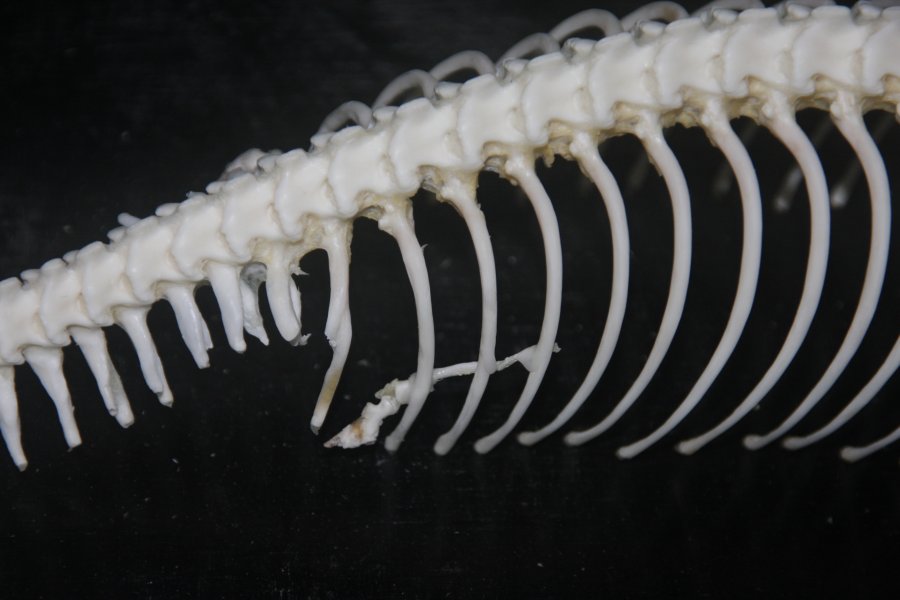
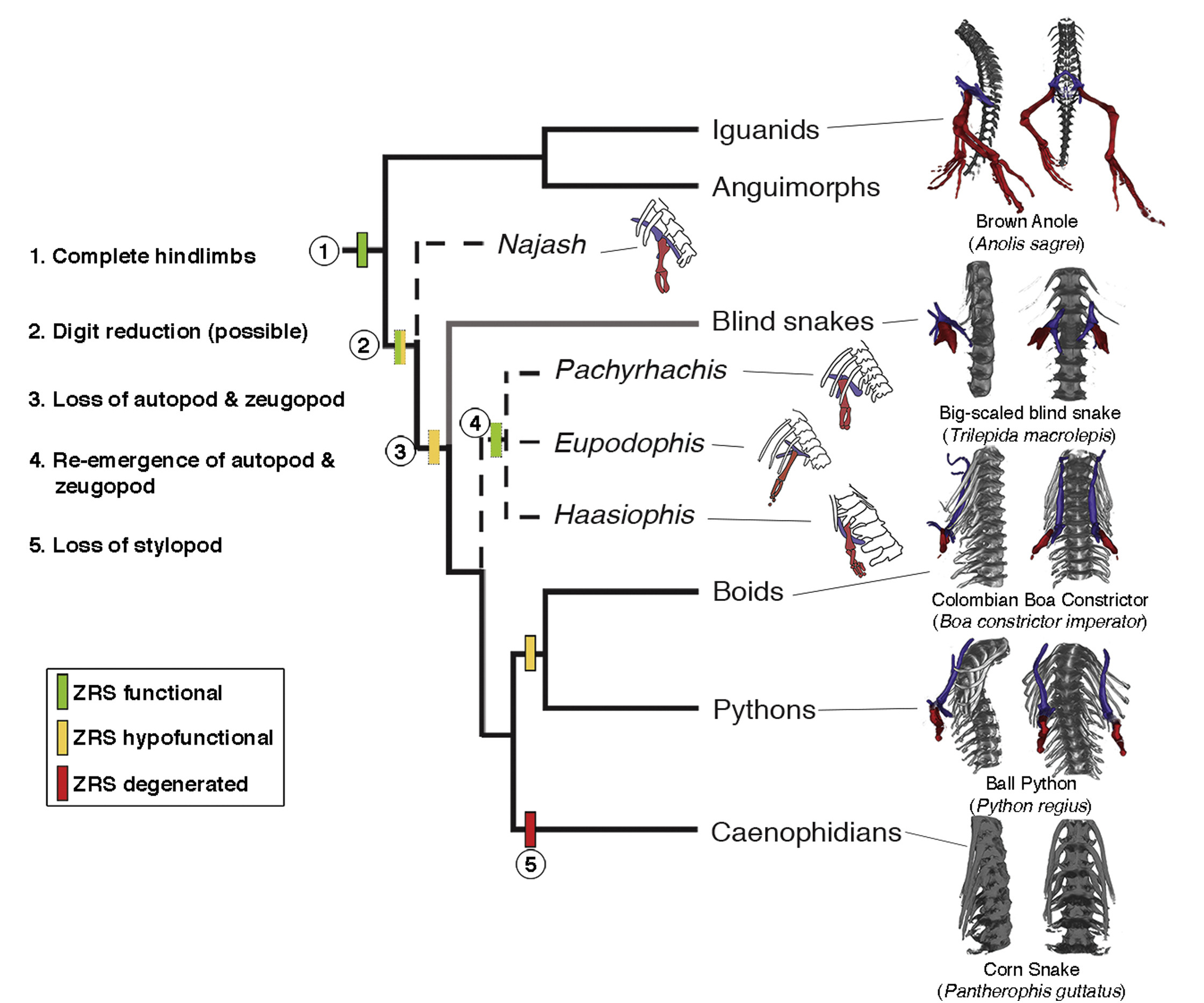
A korábbi posztban emlegetett végtag-specifikus Sonic hedgehog (Shh) gén enhanszer, a ZRS működésképtelenné válása ugyanis nem egyszerre, hanem valószínűleg több lépésben történhetett, erről tanúskodnak a különböző fosszíliák is, de főként az a tény, hogy egyes ősibb kígyófajokban, pl. pitonokban és boákban máig fellelhető egy rendkívül csökevényes, de egyértelműen beazonosítható hátsó végtagcsökevény.
Márpedig ennek a csökevényszervnek a jelenléte arról tanúskodik, hogy a végtagnak a fejlődése elindul, csak valamiért nem zajlik le teljesen. Vagyis logikus kérdés, hogy mi történik pl. a pitonembriók hátsó végtagbimbójában a végtagfejlődésben fontos Shh, vagy az ez által is szabályozott HoxD13 gének expressziójával?
Martin Cohn csoportja erre kereste a választ és így derülhetett fény arra, hogy mindkét gén kifejeződik egy bizonyos szinten, bár a Shh expressziója csak tranziens, a HoxD13 pedig ennek megfelelően sokkal kisebb, mint mondjuk anolisz gyíkokban. A tranziens Shh expresszió azonban nem csak más gének expresszióját befolyásolja a jelek szerint, de jó eséllyel a végtagbimbó sejtjeinek túlélését is: egy lysotracker nevű anyag segítségével jól látható, hogy a kígyók végtagbimbójában a sejtek intenzív pusztulásba kezdenek (ami már önmagában is jó magyarázat a sorvadt végtagra).
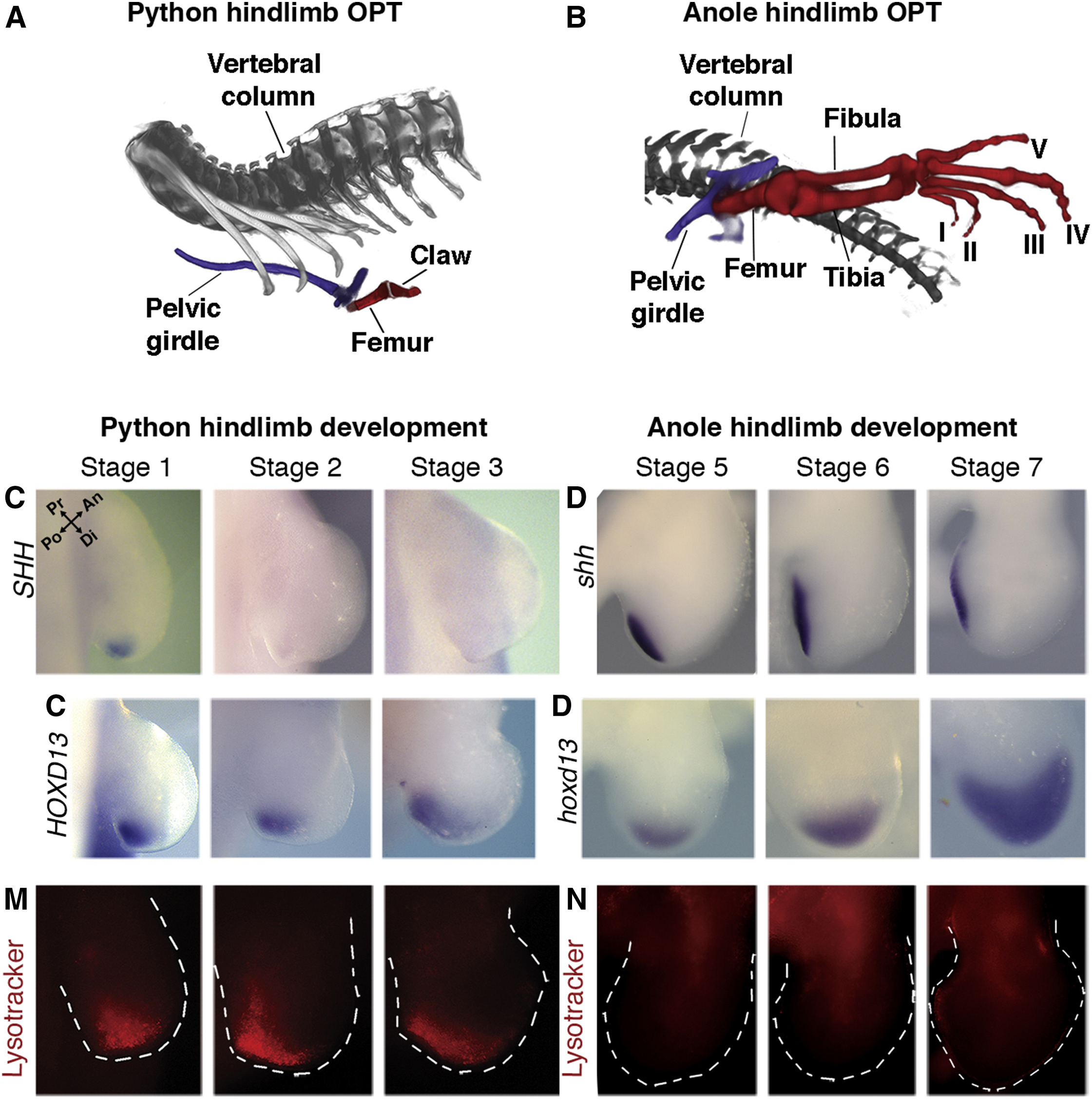
A tanulmány további részében a piton ZRS-ben fellelhető három deléciót vették szemügyre (ezek közül az egyik azonos azzal a 17 bp-al, amit a közelmúltbeli Cell cikk is kielemez), amelyek feltehetőleg fokozatosan jöttek létre, és együttesen hozzák létre a legnagyobb expressziós csökkenést, ha valamilyen riportergén mellett vizsgáljuk a hatásukat.
Mivel a modernebb kígyók esetében a ZRS még inkább elsorvad, az is egyértelművé válik, hogy a pitonok hipofunkcionális ZRS-e végül aztán teljesen elveszti funkcióját. De az érdekes kérdés, hogy a hipofunkcionális ZRS időnként „visszaerősödhetett”-e? A kígyók ma feltételezett filogenetikai leszármazása, ahol a vak fonálkígyók csoportja ősibbnek tekinthető, mint olyan jól felismerhető hátsó végtaggal rendelkező fosszíliák, mint a Haasiophis azt sugallja, hogy vagy a ZRS végső degradálódása többször, függetlenül bekövetkezett, vagy a közös ősben ugyan már a mai pitonokéhoz hasonlóan hipofunkcionális volt, de később egyes, mára kihalt csoportokban a ZRS újból működőképessé vált. (Szerintem utóbbinak van kisebb esélye, de a cikk írói mégis ezt favorizálják.)
A cikk még egy érdekes dologra hívja fel a figyelmet: sokat tudunk a végtagfejlődés során azokról a szabályozó szekvenciákról, amelyek az ujjak fejlődése során szabályozzák a HoxD13 gén működését. Bár végtagok hiányában a pitonok (és más kígyók) esetében ezek látszólag fölöslegesek lettek, mégsem degenerálódtak, hanem teljes egészükben ott pompáznak a kígyógenomokban is. Mi magyarázhatja ezt? A legvalószínűbb magyarázat erre, hogy ezeknek a szekvenciáknak van egy másik nem elhanyagolható szerepe, mégpedig a hím nemiszerv fejlődésében. És erről, még a kígyók sem mondtak le.
Leal F, Cohn MJ (2016) Loss and Re-emergence of Legs in Snakes by Modular Evolution of Sonic hedgehog and HOXD Enhancers. Curr Biol 26(21): 2966-2973.
 Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.
Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.
A denevérek szárnyának kialakulásával már többször is foglalkoztunk itt a blogban, de míg az eddigi tanulmányok leginkább korábbi fejlődéstani ismereteink alapján keresték azokat a géneket, amelyeknek fontos szerepe lehet a szárny létrejöttében, addig a napokban megjelent tanulmány-páros, az újgenerációs szekvenálási technikáknak köszönhetően egy kicsit „elfogulatlanabb”, előfeltételezéseket nélkülöző megközelítést alkalmazott.
Korábban már fény derült arra, hogy az úgynevezett BMP-jelátviteli útvonal különböző tagjai miként járulnak hozzá a kéz csontjainak megnyúlásához, illetve hogyan biztosítják az ujjpercek közti bőrred fennmaradását.
Most, a Nature Genetics-ben megjelent cikk egyrészt a denevér genom fehasználásával összeveti a mellső és hátsó végtag fejlődése során felfedezhető génexpressziós, ill. epigenetikai különbségeket, másrészt beazanosít olyan genomi régiókat, amelyek a denevérek evolúciója során különösen gyorsan változtak – s így feltehető, hogy közük lehet a csoport jellegzetes morfológiai újításának, a szárnynak a kialakításában. Ezek az ún. Bat Accelerated Regions (BAR) szekvenciák jelentik aztán egy PLOS Genetics cikk alapanyagát, és ezekkel fogunk mi is jobban foglalkozni.
 Vannak témák, amelyek végigkövetik a blog most már évtizedes történetét, és az egyik legrégebbi történet, aminek szálait göngyölítgetjük, arról szól, hogy az élővilág evolúciója során miként alakult ki az élőlények formagazdagsága.
Vannak témák, amelyek végigkövetik a blog most már évtizedes történetét, és az egyik legrégebbi történet, aminek szálait göngyölítgetjük, arról szól, hogy az élővilág evolúciója során miként alakult ki az élőlények formagazdagsága.
A sztori megkerülhetetlen eleme a testfelépítésért felelős, ún. Hox gének története épp ezért ha van téma, amivel sokszor, sokat foglalkoztunk, ez biztos az. De néha a „sokat” sem elég, főleg, hogy most már jó ideje egyáltalán, szóval időszerű egy kicsit elmerülni ismét abban a megunhatatlan témában, hogy miként formálta és formálja ez az ősi géncsalád például az ízeltlábúak formáit.
Az „alapsztorit” már bő nyolc és fél éve megírtam, és zanzásítva valahogy úgy szól, hogy az ízeltlábúak jól látható szelvényes, ha úgy tetszik „moduláris” felépítését a Hox gének változó expressziós tartománya befolyásolja. Az ízeltlábúak egyes szelvényeinek alapfelépítése tulajdonképpen nagyon egyszerűen megfogalmazható: egy henger alakú testből egy pár nyúlvány lóg ki. Hogy ez a nyúlvány járólábként, szájszervként, vagy csápként, az a szelvény pozíciójától függ. És ezt a pozicionális információt a Hox gének határozzák meg, amelyek ha elromlanak, a szelvény identitása is elromlik, így szájszervek, vagy csápok helyett péládul járóábak jelennek meg a fej szelvényein is (ezeket nevezzük homeotikus mutációknak).
Amikor aztán sokan azt is megfigyelték, hogy az egyes fontosabb Hox gének kifejeződése nagyobb taxonómiai csoportonként jellegzetes változásokat hordoz, sokan elkezdték fejtegetni, hogy valószínűnek tűnik, sok esetben a csoportok sztereotipikus külalakjában megfigyelhető jellegezetes különbségek is az ilyen Hox-expresszióban jelentkező különbségekre vezethetők vissza. Magyarán az evolúció szívesen „játszott” ezekkel a génekkel.
 A Hox-gének hatását az ízeltlábúak végtag-evolúciójára már hosszú ideje dokumentáljuk a blogon. Mára már kvázi tankönyvi adat lett, hogy ezeknek a homeotikus géneknek az expresszióbeli változásai nagyban befolyásolták, hogy az egyes szelvényeken milyen végtagok jöjjenek létre (pl. rákok szájszervei).
A Hox-gének hatását az ízeltlábúak végtag-evolúciójára már hosszú ideje dokumentáljuk a blogon. Mára már kvázi tankönyvi adat lett, hogy ezeknek a homeotikus géneknek az expresszióbeli változásai nagyban befolyásolták, hogy az egyes szelvényeken milyen végtagok jöjjenek létre (pl. rákok szájszervei).
A legutóbbi példáink a méhek dolgozóinak pollenkosara, a vízipoloskák láhossza és a tücskök ugrólábai voltak. Most egy újabb csoport, az ugróvillások adódnak az eddigi népes példákhoz.
Michael Akam csoportja az Orcheselle cincta nevű ugróvillás fejlődését tanulmányozta, pontosabban azt, hogy miként fejeződik ki az ízeltlábúakban legfontosabb két poszterior Hox-gén, és mi a következménye, ha nem működnek.
 A béka hátsó lábában, a gekkók melső végtagjában és a mi kezünkben a számtalan homológ csontocska mellett is feltünő egy, fontos, közös vonás: mindannyiunknak öt ujja van. Az öt ujjúság a szárazföldi négylábú gerincesek nagy közös vonása, olyannyira, hogy egy tudományos kifejezés is születtet a sztereotip négylábú végtag leírására: pentadactyl végtag.
A béka hátsó lábában, a gekkók melső végtagjában és a mi kezünkben a számtalan homológ csontocska mellett is feltünő egy, fontos, közös vonás: mindannyiunknak öt ujja van. Az öt ujjúság a szárazföldi négylábú gerincesek nagy közös vonása, olyannyira, hogy egy tudományos kifejezés is születtet a sztereotip négylábú végtag leírására: pentadactyl végtag.
Az ettől való eltérés mindig különös és az evolúciósan csak egy irányba történik: a békák melső lábán, a madarak szárnyában, vagy a lovak patájában egyaránt kevesebb ujj figyelhető meg (négy, három, illetve egy). Az ötnél több ujjúság (az ún. polydactylia) kóros esetekben észlelhető, például a végtagok fejlődése apropóján korábban már emlegetett Sonic-hedgehog (Shh) gén mutációja okozhat ilyen elváltozást.
De vajon miért pont öt a normális, illetve lehette-e valamikor hat, vagy hét? Esetleg tizenkettő?
 Amikor egyszer régen írtam egy hosszú, bevezető posztot az emlős végtagok fejlődéséről, akkor az volt a nem is igen titkolt szándékom, hogy az egész amolyan felvezetésként szolgál majd a mellső végtag evolúcióját tárgyaló posztokhoz. Aztán teltek az évek és igazából csak nem sikerült időt szakítani a témára, pedig mindig lett volna anyag. Múlt hónapban azonban két olyan fontos cikk is kijött, ami miatt most mégis össze kell kapnom magam, nem is egy, hanem két poszt erejéig, mert ezekről hallgatni már-már bűn lenne. 🙂
Amikor egyszer régen írtam egy hosszú, bevezető posztot az emlős végtagok fejlődéséről, akkor az volt a nem is igen titkolt szándékom, hogy az egész amolyan felvezetésként szolgál majd a mellső végtag evolúcióját tárgyaló posztokhoz. Aztán teltek az évek és igazából csak nem sikerült időt szakítani a témára, pedig mindig lett volna anyag. Múlt hónapban azonban két olyan fontos cikk is kijött, ami miatt most mégis össze kell kapnom magam, nem is egy, hanem két poszt erejéig, mert ezekről hallgatni már-már bűn lenne. 🙂
Az első cikk azzal foglalkozik, hogy miképpen befolyásol(hat)ta az elnyújtott Hoxd13 expresszió a végtagok fejlődését. De, hogy ne rontsunk ajtóstól a házba, előbb lássuk, hogyan kerül elő egyáltalán ez a gén és egyáltalán, miben is különbözik egy egér és egy hal végtagja.
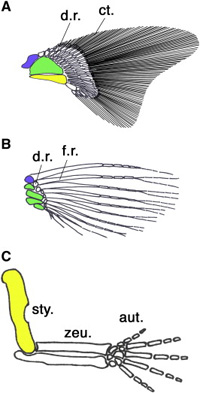
Ha az ősi gerinces végtagot vesszük szemügyre, ami ma is megfigyelhető egyes procoshalakban (lásd cápák – A), akkor azt látjuk, hogy a végtag ízesülését a vállövhöz három nagy csont biztosítja, ezek a propterygium (kék), mesopterygium (zöld) és metapterygium (sárga). Ezekhez apróbb csontok kapcsolódnak (distal radials – d.r.), amelyek az úszósugarakat tartják. Csontoshalak esetében (B) ez a szabásminta úgy egyszerűsödött, hogy a metapterygium elvesződött, és az úszósugarakat tartó apróbb csontok száma is redukálódott. A szárazföldi gerincesekben (C) viszont pont a metapterygium maradt egyedül meg (felkarcsontként), a disztális radiálisokból pedig kialakultak az alkarunk csontjai és a kéztőcsontok. (Némi nemű vita van arról, hogy az ujjak maguk az úszósugarak homológjai-e, vagy sem, de a jelenleg dominánsabbnak tűnő nézet szerint igen.)
 A méhek kapcsán az utóbbi időben kicsit a méhcsaládösszeomlás enigmájára koncentrállgattunk errefele, de most megtörném a nemes hagyományt és egy másik nagyon érdekes aspektusát boncolgatnám a méhek biológiájának, mégpedig a lábak morfológiáját.
A méhek kapcsán az utóbbi időben kicsit a méhcsaládösszeomlás enigmájára koncentrállgattunk errefele, de most megtörném a nemes hagyományt és egy másik nagyon érdekes aspektusát boncolgatnám a méhek biológiájának, mégpedig a lábak morfológiáját.
Kicsit pontosítva, a hátsó lábakról lenne szó és még pontosabban azok ivari kétféleségéről, azaz dimorfimzmusáról, hogy miért olyanok, amilyenek egy dolgozóban és egy királynőben.
Mielőtt azonban ennyire előre rohannánk, előbb egy picit ismerkedjünk meg a szóban forgó testrésszel. A méhek lába tipikus rovarokra jellemző ízeltláb, ennek megfelelően részei a lábtő (coxa), a tompor (trochanter), comb (femur), lábszár (tibia), illetve a lábat alkotó basitarsus és tarsus.
A méhek esetében hátsó (harmadik) pár lábon a lábszáron egy különleges képződmény jelenik meg, az ún. pollen kosár (pollen basket). Ennek funkciója, mint arra neve is utal, a pollen elszállításában van: a rajta levő nagyméretű szőrszálak kifejezetten alklamasak a ragacsos pollen megtartására, így a dolgozók nagyméretű pollengalacsinokat képesek hazaszállítani segítségükkel.
 Homebox géneket szeretni már-már klisé szinten sztereotipikus egy fejlődésbiológustól, ugyanakkor tény, hogy a leglátványosabb mutációkért és evolúciós változásokért felelős géncsaládot nem lehet nem elismerni. Ennek megfelelően időről-időre mi is lerovunk néhány tiszetelkört a batesoni homeotikus mutációk genetikai mibenlétét biztosító DNS szakaszok előtt.
Homebox géneket szeretni már-már klisé szinten sztereotipikus egy fejlődésbiológustól, ugyanakkor tény, hogy a leglátványosabb mutációkért és evolúciós változásokért felelős géncsaládot nem lehet nem elismerni. Ennek megfelelően időről-időre mi is lerovunk néhány tiszetelkört a batesoni homeotikus mutációk genetikai mibenlétét biztosító DNS szakaszok előtt.
A Hox gének külön érdekes szerepet játszottak az ízeltlábúak evolúciójában, ahol a szájszervek alakja, a szárnyak helyzete és a potroh láb-nélkülivé válása egyaránt nekik köszönhető.
Ugyanakkor, paradox módon az utóbbiak esetében (azaz a potroh végtag-mentességét biztosító poszterior Hox gének, az ubx és abdA esetében) az utóbbi évek egy új, a végtagok hosszát befolyásoló funkiót is feltártak.

A púposkabócák (Membracidae) hátát díszítő gyakran szürreális képletek bárki érdeklődését könnyen felkeltik. A részben szexuális szelekció, részben mimikri következtében kialakult kacifántos képződményeket eddig az első torszelvény nyúlványainak tekintették, de mint arra most fény derült, eredetük sokkal izgalmasabb.
Sean Carroll két volt tanítványa, Benjamin Prud’homme és Nick Gompel vezetett csoport ugyanis nem kevesebbet bizonyított be, mint, hogy valójában módosult szárnyakról van szó.