Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.
A denevérek szárnyának kialakulásával már többször is foglalkoztunk itt a blogban, de míg az eddigi tanulmányok leginkább korábbi fejlődéstani ismereteink alapján keresték azokat a géneket, amelyeknek fontos szerepe lehet a szárny létrejöttében, addig a napokban megjelent tanulmány-páros, az újgenerációs szekvenálási technikáknak köszönhetően egy kicsit „elfogulatlanabb”, előfeltételezéseket nélkülöző megközelítést alkalmazott.
Korábban már fény derült arra, hogy az úgynevezett BMP-jelátviteli útvonal különböző tagjai miként járulnak hozzá a kéz csontjainak megnyúlásához, illetve hogyan biztosítják az ujjpercek közti bőrred fennmaradását.
Most, a Nature Genetics-ben megjelent cikk egyrészt a denevér genom fehasználásával összeveti a mellső és hátsó végtag fejlődése során felfedezhető génexpressziós, ill. epigenetikai különbségeket, másrészt beazanosít olyan genomi régiókat, amelyek a denevérek evolúciója során különösen gyorsan változtak – s így feltehető, hogy közük lehet a csoport jellegzetes morfológiai újításának, a szárnynak a kialakításában. Ezek az ún. Bat Accelerated Regions (BAR) szekvenciák jelentik aztán egy PLOS Genetics cikk alapanyagát, és ezekkel fogunk mi is jobban foglalkozni.
Darwin óta a biológia egyik központi kérdése, hogy apró, fokozatos változások során hogyan jönnek létre a nagy küllembeli különbségek. És ugyan az ún. „hopeful-monster” elmélet (amely szerint a nagy és drámai változások hirtelen jelentek meg, kvázi egyetlen mutáció révén) időről időre felüti a fejét (most épp a Jerry Coyne ostorozza az elmélet híveit a The Loom-on), mára a többségi konszenzus a gradualitás mellett van.
Az ilyen drámai változások vizsgálatában az talán a legnehezebb, hogy a két fajt nem keresztezhetjük egyszerűen, hogy aztán klasszikus genetikai térképezés során megkereshessük a változás genetikai okait. Marad tehát az ún. „candidate approach” , vagyis olyan géneket veszünk szemügyre, amelyekről tudjuk, hogy a két faj egyikében szerepet játszik a szóbanforgó testtáj fejlődésében. S hogy hagyjuk az elvont általánosításokat , lássunk egy konkrét példát: a denevérszárny kialakulását.
Ha összehasonlítjuk a denevérek és az egerek mellső végtagját, a bőrredők mellett a legfeltűnőbb különbség, hogy a denevérek „keze” sokkal hosszabb. Mivel egerek (és emberek) esetében a csontok fejlődésében egy Bone Morphgenetic Factor (BMP) fedőnevű molekulacsalád kiemelt szerepet játszik, a „candidate approach” keretében természetesen erősen ezekre a fehérjékre koncentráltak. Az eredmény egyrészt igazolta a módszer hatékonyságát, hiszen a denevérek mellső végtagjaiban valóban jelentősen megnőtt a BMP-khez kapcsolható jelátviteli útvonalak aktivitása, másrészt arra nem adott konkrét választ arra kérdésre, hogy pontosan mi okozta ezt a növekedést. A BMP-kódoló génekben következett be valami mutáció, vagy az őket is szabályozó transzkripciós faktorok valamelyikében.
Ezt eldöntendő, egy amerikai csoport a BMP-szabályozó képességéről ismert Prx1 transzkripciós faktort kezdte vizsgálni. A Prx1 jó helyen és időben fejeződik ki a végtagokban: pont a növekedési fázis során, a növekedésért felelős, központi régió közelében. Ráadásul, ha hiányzik, az egerek keze-lába jelentősen rövidebb lesz.
Először a Prx1 denevér megfelelőjét különítették el, megvizsgálva a genomi szerkezetét és a kifejeződési mintázatát. Utóbbi igencsak ígéretesnek mutatkozott, mert nemcsak hogy jelen volt a fejlődő szárnyban, de lényegesen szélesebb tartományt fedett le, mint egerekben. Következő lépésben izoláltak egy-egy olyan szabályozó szekvenciát (enhancert), amelyik egerekben illetve denevérekben a gén kifejeződésének pontos helyét regulálják, és az egér enhancert kicserélték a denevér-specifikus enhancerre.
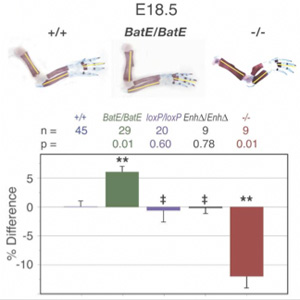
Az így létrehozott egerek (BatE/BatE) mellső végtagja hosszabnak bizonyult a normális (+/+) egérkarnál (hogy a már említett Prx1 mutánsokról (-/-) ne is szóljunk), és ez a változás azzal volt összefüggésben, hogy fejlődésük során a csontok több osztódó sejtet tartalmaztak – épp mint a denevér-mellsővégtag esetében. Vagyis, a jelek szerint Prx1 denevér specifikus enhancere tartalmazza az egyik (de naná, hogy messze nem az egyetlen) olyan apró változást, ami felelőssé tehető a szárny és a mellső mancs közötti alaktani különbségért.
Az egyetlen meglepetés akkor érte a kutatókat, amikor kitörölték az egérs-specifikus Prx1 enhancert, de nem ültettek semmit a helyére. Intuitív módon azt várná az ember, hogy a Prx1-/- mutánshoz hasonlóan, ez a genetikai változás is rövidebb végtagokat eredményez, de nem ez történt: ezek az egerek (EnhΔ/EnhΔ) pont ugyanakkora végtaggal rendelkeztek, mint nemmutáns társaik. A jelenség pontos oka nem ismert, de a legvalószínűbb magyarázatnak az tűnik, hogy a kutatók által izolált enhancer legalábbis az egerekben „redundáns”, vagyis más enhancerek is ellátnak hasonló funkciót, így azok kárpótolni tudták a kieső szabályozó szekvenciát a mutánsokban. Hogy ez igaz vagy sem, az majd akkor válik el, ha megtalálják a további Prx1 enhancereket.
Cooper KL, Tabin CJ (2008) Understanding of bat wing evolution takes flight. Genes Dev22(2): 121-124. Cretekos CJ, Wang Y, Green ED; NISC Comparative Sequencing Program, Martin JF et al. (2008) Regulatory divergence modifies limb length between mammals. Genes Dev22(2): 141-151.
Ahogy egy-egy szó jelentését is nagymértékben befolyásolja a szövegkörnyezet, úgy az egyes fehérjék „jelentését”, funkcióját is nagymértékben befolyásolja, hogy milyen molekuláris milliőbe kerülnek, milyen más fehérjékkel kerülnek kölcsönhatásba.
A BMP fehérjék visszatérő vendégeink, és már korábban többször is szóltam arról, hogy az egyik legkarakterisztikusabb tulajdonságuk, hogy elősegítik a csontnövekedést – pl. a denevérszárny ujjperceinek meghosszabbodását is. De a BMP-knek tucatnyi más feladatuk is van az egyedfejlődés során, s ezek gyakran térben és időben szinte átfednek a korábban már említett feladatokkal (a funkcionális különbség a molekuláris partnerek másságából adódik).
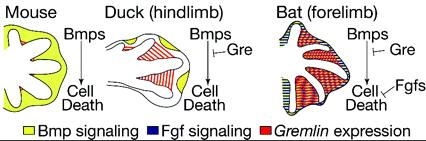
Ennek egészen jó példája a denevérszárnyak esete (hogy témánál maradjunk), ahol az ujjpercek porcosodását és megnyúlását megelőzően a bmp expresszió pont az ujjak közötti szövetekben figyelhető meg, ahol a szóbanforgó sejtek elpusztulását idézi elő. Ezért nem lesz aztán semmilyen hártya az ujjaink között. Mármint nekünk, mert más fajok esetében van erre példa. Pl. a kacsák esetében, ahol elég pontos képünk van arról, hogy miként gátolódik a BMP-függő sejthalál az ujjközti területeken. Egyrészt a BMP jelátvitel egyik effektora, az msx gén nem fejeződik ki az említett régióban (ellentétben pl. a csirkékkel, vagy egerekkel), másrészt pedig megjelenik a gremlin nevű gén terméke, amely egy igen potens BMP anatgonista.
Most néhányan annak eredtek a nyomába, hogy mennyire hasonló okok miatt marad meg a hártya a denevér ujjai között. Mint kiderült, még a konvergens evolúció sem lép kétszer tökéletesen ugyanabba a folyóba: az msx expresszió a denevér embrióban gyakorlatilag megegyezik az egérembrióban megfigyeltekkel (vagyis ez nem lehet az oka az apoptózis elmaradásának). Viszont a gremlin expresszió valóban megjelenik a denevérek mellső végtagkezdeményében. S bár ez már önmagában is elegendő lehetne a sejtpusztulás elmaradásának magyarázatra, a jelek szerint működik még egy párhuzamos mechanizmus is: egy fibroblast growth factor (fgf) gén is elkezd kifejeződni a leendő hártyában, ami szintén a sejthalál gátlásához vezet.
Summa-summarum, a hártyás szárny (vagy éppen láb) nem a semmiből jelent meg az evolúció során; mindig is jelen volt a négylábú gerinces embriók fejlődésének egy adott szakaszában. S amikor az élőlénynek haszna számazhatott abból, ha ez a hártya megmarad, a szelekció révén kialakultak azok a molekuláris „fékek”, amelyek meggátolták elhalását.
Weatherbee, SD, Behringer, RR, Rasweiler, JJ, Niswander, LA (2006) Interdigital webbing retention in bat wings illustrates genetic changes underlying amniote limb diversification. PNAS103: 15103-15107.
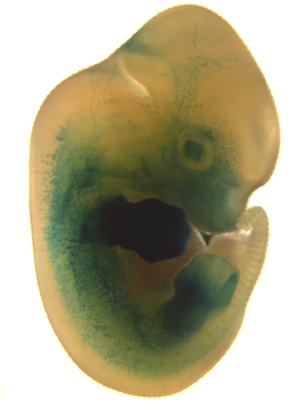
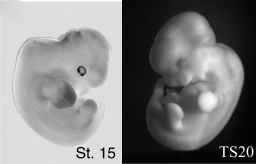
Az élővilág változatosságának legfőbb oka az embrionális fejlődés során bekövetkező kisebb-nagyobb változásokban rejlik. Általában minnél közelebbi rokon két faj, ezek a változások annál később válnak nyilvánvalóvá az egyedfejlődés során. Leggyakrabban nincs is szó új gének megjelenéséről, egyszerűen meglevő gének szabályozása változik meg egy kicsit. Mint a mellékelt ábrán látható pl. az embrionális fejlődés egészen késői szakaszáig a denevér fejlődése (bal oldalon) kísértetiesen hasonlít az egér fejlődésére (jobb oldalon). Már jól elkülönülnek az egyes testtájak, de még nyoma sincs a későbbi szárnynak és hegyes füleknek. Ezek később válnak láthatóvá, de a hozzájuk vezető molekuláris változások az ábrán látható idő tájékán következnek be.
A denevérek alakja nem új keletű, már az 50 millió éves fosszíliák (fent) a ma élő utódiakhoz (lent) hasonló szárnystruktúrával rendelkeztek. Ez pedig azt jelenti, hogy a hüvelykujjuk kivételével ujjperceik meghosszabodtak, s a köztük feszülő bőrredő révén úgynevezett „kéz-szárnyat” alkotnak. (Ezzel szemben a madarak szárnya ún. „kar-szárny” – amely a teljes kar módosulása révén alakult ki -, a Pterosaurusok szárnya pedig „ujj-szárny” volt – itt egyetlen ujj részeinek a meghosszabbodása képezte a szárny alapját.) Csak erre az alakra nézve, világossá válik, hogy a szárnykeletkezés kezdeti lépéseit valahol a mellső végatg fejlődése során kell keresni.
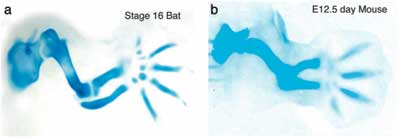
A végtagbimbók fejlődése és növekedése során fokozatosan alakulnak ki a kart alkotó csontok. Először sejtek vándorlása és gyülekezése révén egy porc alapú „váz” jön létre, amelyet később fokozatosan helyettesít a csontszövet. Ez az a pont ahol az egér és a denevér végtagjainak kialakulását figyelve egy fontos különbséget lehet észrevenni: utóbbiak esetében a kialakuló ujjak porcos váza nem áll meg a növekedésben.
A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd egyes halak és pintyek arcberendezésének alakulását), de végtagok esetén a tudományos irodalom további tippekkel is szolgál. Ha egerek végtagbimbóit Bmp2 gazdag médiumban nevelgetjük: az ujjak hosszabra nőnek, ellenben, ha egy Bmp anatgonistát, Noggint keverünk a tápoldatba, az tompa, rövid ujjakhoz jutunk. Ezen a gondolati úton végighaladva nem nehéz kitalálni, mi is az első teendő, ha a denevér ujjak megnyúlásának okát keressük: megvizsgálni, mi történik a kézben kifejeződő bmp2 génnel.
A fentiek ismeretében az eredmény nem teljesen meglepő: a denevérek fejlődő mellső végtagjának ujjaiban mind a bmp2 expresszió, mind a Bmp-k által szabályozott Smad jelátviteli út aktivitása sokszorosa annak, amit a hátsóvégtagjukban, vagy egerek mellső végtagjában figyelhetünk meg. Azaz a szárnykialakulás egy fontos (bár nem kizárólagos) tényezője a szóbanforgó gén aktivitásának felerősödése lehetett egy olyan helyen, ahol amúgy is kifejeződött az egyedfejlődés során.
A denevérek mellső végtagja fejlődő ujjperceinek sejtjeiben (kék) lényegesen magassabb a Bmp fehérjék (piros) szintje, min a hátsó végatag ujjperceiben, vagy az egerek mellső végtagjában (e-g). Mindez a bmp2 gén magasabb aktivitásával függ össze (h) és ennek következményeként a Smad jelátviteli út is sokkal több sejtben kerül aktiválásra (piros) a denevér kéz ujjaiban mint lábukban vagy az egerekben (i-k).
Azok pedig akik azt szeretnék tudni, hogy a későbbiekben mi történik a denevér embrióval, a Pharyngulán elégíthetik ki kiváncsiságukat. 😉
Sears KE, Behringer RR, Rasweiler JJ, Niswander LA (2006) Development of bat flight: Morphologic and molecular evolution of bat wing digits. Proc. Nat. Acad. Sci. USA103(17): 6581-6586.

 Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.
Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.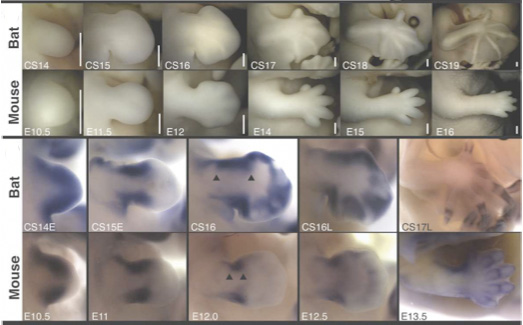


 A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd
A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd 