
Mint arra a legelső blogbejegyzésben utaltam, az élőlények alakjaiban bekövetkező változások általában egyes gének felhasználási helyének és idejének változására vezethetők vissza, s ezen változások képezik az evolúció egyik hajtóerejét. Az ilyen típusú változásokat (értelemszerűen) leginkább olyan rokon fajokkal lehet szemléltetni melyek evolúciós időskálán mérve a közelmúltban váltak el, és ha ezen rokon fajok nagyfokú adaptív radiáció (igen különböző ökológiai életterekhez, ún. nichekhez való alkalmazkodás) folytán jöttek létre az érv általában még meggyőzőbb erejű.
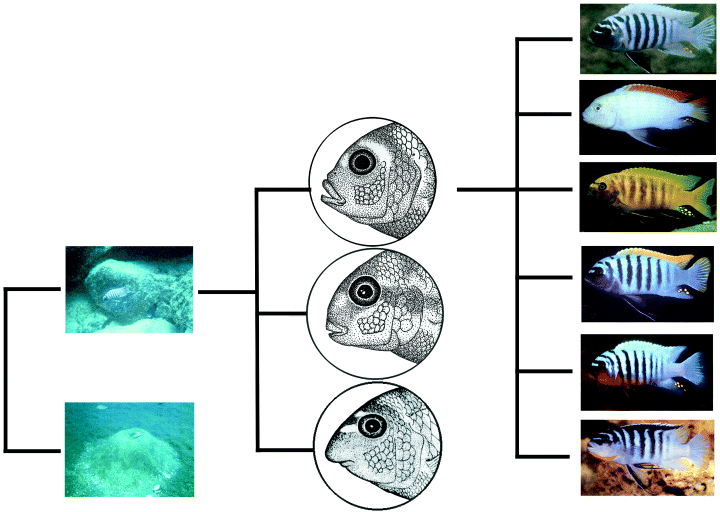
1. Ábra A bölcsőszájú halak adaptív radiációjának legfontosabb lépései (először élőhelyhez, majd táplálékforráshoz való alklamazkodás, végül szexuális szelekció). [1]
A fenti állítás szemléltetésére talán az egyik legtriviálisabb példa a bölcsőszájú halak esete. Kelet-Afrika Nagy Tavaiban (Tanganyika, Viktória, Malawi) mintegy 400 bölcsőszájú halfaj él, melyek egy közös ősből alakultak ki cirka 700.000 év leforgása alatt. Feltehetőleg három nagy adaptációs „robbanás” játszott szerepet a fajok kialakulásában (1. ábra). Először az élőhely alapján két csoport különült el: a homokos aljzat közelében illetve a sziklás környezetben lakók, majd utóbbiak a táplálkozási szokásuk alapján tovább specializálódtak szilárd tápanyagot rágcsálókra illetve lebegő tápanyagot fogyasztókra. Végül szexuális szelekció során jöttek létre a különböző színes mintázatok [1].
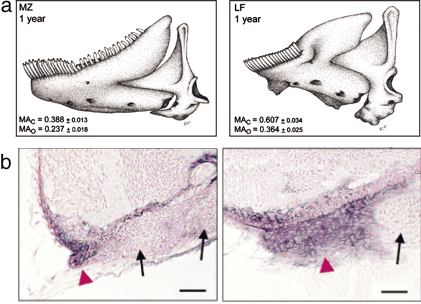 2. Ábra A M. zebra karcsúbb állkapcsának kialakulása összefügg azzal, hogy bmp4 mRNS csak az első kopoltyúív végében található, ellentétben a L. fuellerbonival, ahol a masszív állkapocs az egész kopoltyúívben jelenlevő bmp4-el társul (készült [2] alapján).
2. Ábra A M. zebra karcsúbb állkapcsának kialakulása összefügg azzal, hogy bmp4 mRNS csak az első kopoltyúív végében található, ellentétben a L. fuellerbonival, ahol a masszív állkapocs az egész kopoltyúívben jelenlevő bmp4-el társul (készült [2] alapján).
Egy közelmúltban megjelent tanulmány [2] a második alkalmazkodási kör okait vizsgálja. Ehhez két különböző táplálkozási szokású halfajt hasonlítottak össze, ezek a Labeotropheus fuelleborni (LF) ill. Metriaclima zebra (MZ). Elöbbinek a sziklákon levő algák rágcsálásához alkalmazkodott rövid, de robusztus alsó állkapcsa van, míg utóbbinak karcsúbb és hosszabb az említett testrésze (minthogy a szóbanforgó hal mindenevő, ez nem meglepő). A két fajt keresztezve és a lérejövő ún. F1, illetve az ennek a keresztezéséből származó F2 nemzedék egyedeit vizsgálva lehetőség nyílt annak a meghatározására, hogy melyik kromoszómán vannak az állkapocs méretét/alakját befolyásoló gének. Ez a meghatározás azért lehetséges, mert az utódok nyilvánvalóan nem teljesen egyformák, sem genetikailag, sem külalakjukat tekintve, így kapcsolatot lehet találni bizonyos kromoszómális genetikai markerek és az állkapocs egyes jellegzetes paraméterei között. (Egy fontos megjegyzés: az igazán szemfülesek biztos kiszúrták, hogy két faj keresztezéséről beszélek, ami az állatoknál alkalmazott faj definíció értelmében max. steril utódokat hozhatna létre, így F2 szóba sem jöhet. A helyzet az, hogy a faj definíciója egy mesterséges kategória, és bár általában igaz, nem feltétlenül alkalmazható mindig. Jelen esetben a két faj között a különböző színezet miatt a természetben létezik egyfajta szexuális izoláció, de azért annyira (még) nem különböznek egymástól, hogy ne hozzanak létre életképes hibrideket.) A vizsgálat egyik érdekes eredménye, hogy bizonyos tulajdonságok együtt változnak: az állkapocs hossza, illetve magassága (ínyencek kedvéért: a coronalis processus mérete) között negatív összefüggés van (azaz minnél rövidebb az állkapocs, annál magassabb egyben). A másik pedig, hogy ezek a tulajdonságok (egyebek mellett) a bone morphogenetic protein 4 (bmp4) nevű génhez kapcsolódnak.
A bmp4, mint neve is mutatja, azon faktorok osztályába tartozik, amelyek a csontfejlődésben is szerepet játszanak (még rengeteg más szerepük is van, de a téma szempontjából ez a legfontosabb). Egyebek mellett egyes arccsontokban is expresszálódik, ami érthető módon itt külön érdekessé teszi. Ezek után már nem is annyira neglepő, hogy a vizsgálat tárgyát képező két halfajban jelentősen különbözik a bmp4 átíródásának helye és mértéke. Míg MZ egyedekben az első kopoltyúív (ez hozza létre az alsó állkapcsot) végére korlátozódik a jelenléte, addig a robusztusabb állkapcsú LF halakban az egész kopoltyúívet kitölti (2. Ábra). Eddig szép és jó a dolog, de, hogy kerek legyen a történet okozati összefüggést kell kimutatni a bmp4 expresszió változás és az állkapocs alakja között. Ehhez a jól bejáratott „kísérleti nyulat”, a zebrahalat használták: mesterségesen megnövelve a Bmp4 szintet a halak állkapcsa sokkal robusztusabb lett. (A történet ott sántít egy kicsit, hogy ez a robusztusabb forma nem járt együtt az állkapocs hosszának csökkenésével, ennek egyik valószínű oka lehet, hogy a Bmp4 mennyiségét az összes szövetben megemelték és nemcsak az első kopoltyúívben.
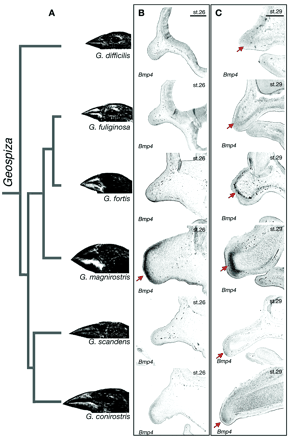 3. Ábra A különböző Geospiza fajok csőralakja összefügg a bmp4 expresszió szintjével. [3]
3. Ábra A különböző Geospiza fajok csőralakja összefügg a bmp4 expresszió szintjével. [3]
És ha már Bmp4 valamint adaptív radiáció, akkor nem mehetünk el a legklasszikusabb példa mellett: Darwin pintyeinek csőrformája.
Ha az evolúcióval kapcsolatban másra nem is igen, de arra majdnem mindenki emlékezni fog, hogy Darwint a Galapagos szigetken elő pintyek (Geospiza fajok) sokfélesége is megihlette elmélete kidolgozásakor. A szigetcsoporthoz tartozó földdarabokon közös őstől származó, de igen változatos életformákhoz alkalmazkodott pintyek éledegélnek és a ez az alkalmazkodás legjobban talán csőrük alakjában nyilvánul meg. A csőrök formagazdagsága mögötti embriológiai okokat keresve egy harvardi csoport végignyálazta azokat a növekedési faktorokat, amelyekről tudott, hogy szerepük van a madár arckoponya kialakulásában. Véletlen vagy sem, de épp a bmp4 volt az amelyik változott a különböző fajok között (3. Ábra), mégpedig úgy, hogy a szélesebb csőrrel rendelkező madarak csőr mesenchymájában megnőtt a szintje [3]. Az okozati összefüggést természetesen itt is vizsgálták és csirkeembriók csőrkezdeményében növelték meg mesterségesen a Bmp4 vagy a Noggin (egy Bmp antagonista) szintet, ami a várt eredményt hozta: előbbi esetben a csőr mérete megnőtt, míg utóbbiban lecsökkent.
Nem teljesen világos, hogy pontosan milyen változásokat okoz a bmp4 expressziós szintjének megemelkedése, de feltehetőleg a sejtek osztódását segíti elő és így növeli a méretet, legalább is erre utal a csirke- és kacsacsőr összevetése [4].
A galapagosi pintyek történetének egyik legszebb mozzanata, hogy nemcsak az evolúció ihletői voltak, de ma is talán az egyik legjobb szemléltetői. Peter és Rosemary Grant, a Princeton egyetem két kutatója hosszú évtizedek óta vizsgálja ezeket a kis madarakat. Ennyi idő alatt szárazabb és nedvesebb periódusok követték egymást a szigeteken, és a pintyek test- valamint csőr mérete ezek függvényében változott. Az 1977-es szárazságot követően a szigeteken csak nagy és kemény magok fordultak elő, aki ezeket nem tudta fogyasztani az éhenhalt. Ennek megfelelően az egyik vizsgált pintyfaj (G. fortis) populációinak csőrmérete módosult és robusztusabb lett, alkalmasabb a kemény magok feltörésére. Néhány évvel később azonban a kedvező esők miatt sok apró mag és kevesebb nagyobb volt fellelhető a szigeteken. Ekkor az előbbiek fogyasztására alkalmasabb kisebb csőr jelentette az evolúciós előnyt. Talán mondanom sem kell, hogy a ezekben az években milyen irányban változott a pintypopuláció csőrmérete [5]… ;-))
[1] Danley PD, Kocher TD (2001) Speciation in rapidly diverging systems: lessons from Lake Malawi. Molecular Ecology 10(5): 1075-1086.
[2] Albertson RC, Streelman JT, Kocher TD, Yelick PC (2005) Integration and evolution of the cichlid mandible: The molecular basis of alternate feeding strategies. Proc.Nat.Acad.Sci. USA 102(45): 16287-16292.
[3] Abzhanov A, Protas M, Grant BR, Grant PR, Tabin CJ. Bmp4 and morphological variation of beaks in Darwin’s finches. Science 305: 1462-1465.
[4] Wu P, Jiang TX, Suksaweang S, Widelitz RB, Chuong CM. Molecular shaping of the beak. Science 305: 1465-1466.
[5] Grant PR, Grant BR. (2002) Unpredictable evolution in a 30-year study of Darwin’s finches. Science; 296: 707-711.