
 Vannak témák, amelyek végigkövetik a blog most már évtizedes történetét, és az egyik legrégebbi történet, aminek szálait göngyölítgetjük, arról szól, hogy az élővilág evolúciója során miként alakult ki az élőlények formagazdagsága.
Vannak témák, amelyek végigkövetik a blog most már évtizedes történetét, és az egyik legrégebbi történet, aminek szálait göngyölítgetjük, arról szól, hogy az élővilág evolúciója során miként alakult ki az élőlények formagazdagsága.
A sztori megkerülhetetlen eleme a testfelépítésért felelős, ún. Hox gének története épp ezért ha van téma, amivel sokszor, sokat foglalkoztunk, ez biztos az. De néha a „sokat” sem elég, főleg, hogy most már jó ideje egyáltalán, szóval időszerű egy kicsit elmerülni ismét abban a megunhatatlan témában, hogy miként formálta és formálja ez az ősi géncsalád például az ízeltlábúak formáit.
Az „alapsztorit” már bő nyolc és fél éve megírtam, és zanzásítva valahogy úgy szól, hogy az ízeltlábúak jól látható szelvényes, ha úgy tetszik „moduláris” felépítését a Hox gének változó expressziós tartománya befolyásolja. Az ízeltlábúak egyes szelvényeinek alapfelépítése tulajdonképpen nagyon egyszerűen megfogalmazható: egy henger alakú testből egy pár nyúlvány lóg ki. Hogy ez a nyúlvány járólábként, szájszervként, vagy csápként, az a szelvény pozíciójától függ. És ezt a pozicionális információt a Hox gének határozzák meg, amelyek ha elromlanak, a szelvény identitása is elromlik, így szájszervek, vagy csápok helyett péládul járóábak jelennek meg a fej szelvényein is (ezeket nevezzük homeotikus mutációknak).
Amikor aztán sokan azt is megfigyelték, hogy az egyes fontosabb Hox gének kifejeződése nagyobb taxonómiai csoportonként jellegzetes változásokat hordoz, sokan elkezdték fejtegetni, hogy valószínűnek tűnik, sok esetben a csoportok sztereotipikus külalakjában megfigyelhető jellegezetes különbségek is az ilyen Hox-expresszióban jelentkező különbségekre vezethetők vissza. Magyarán az evolúció szívesen „játszott” ezekkel a génekkel.
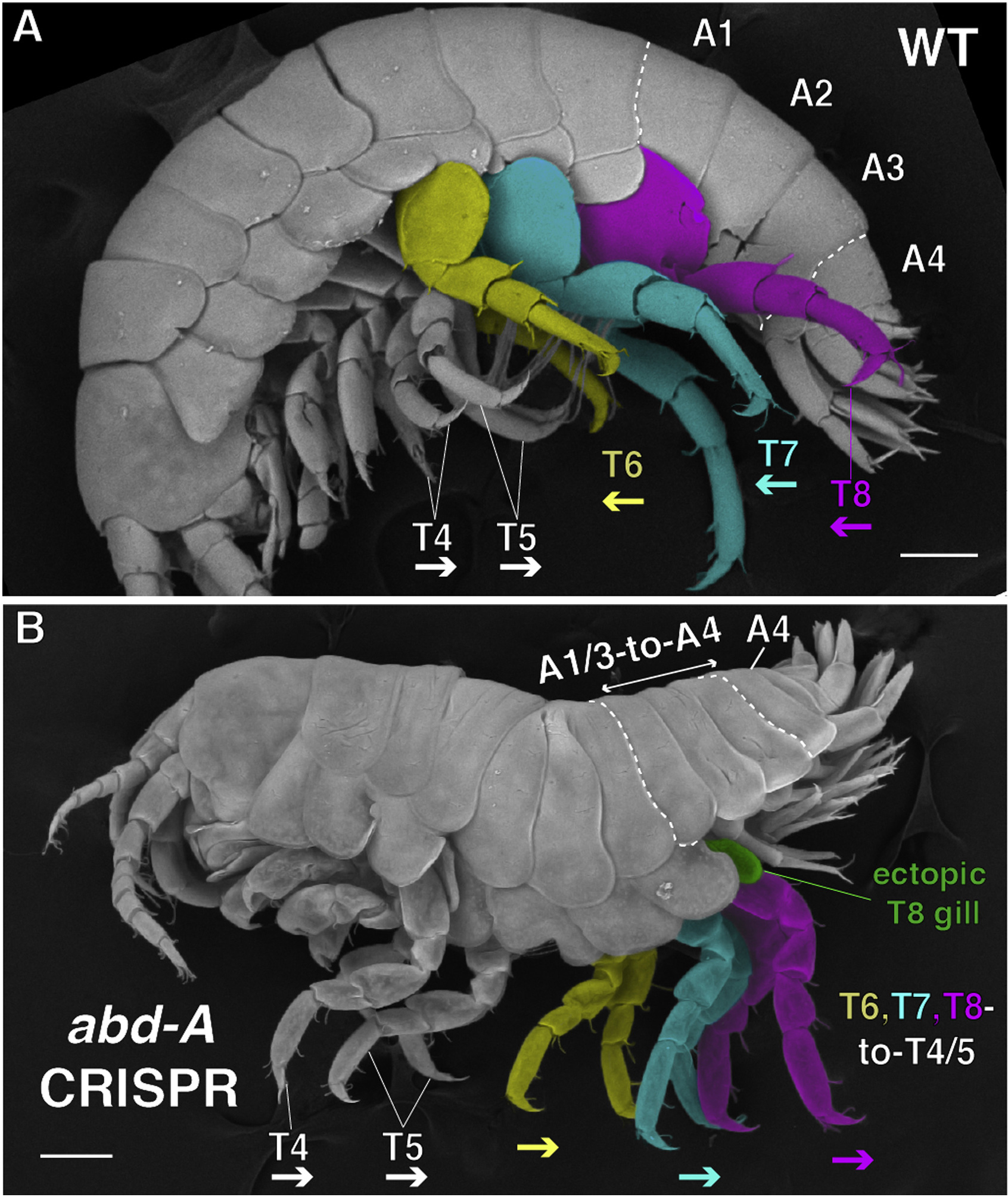
Az egyik immár lassan két évtizedes múltra visszatekintő elmélet szerint az, hogy egyes anterior szelvények nyúlványaiból járó-, vagy állkapcsolábak alakulnak ki, egyetlen gén, az Ubithorax (Ubx) expressziójától függ. Ennek az egyik legelegánsabb bizonyítását 2009-ben az evo-devo kutatás két fontos alakja, Nipam Patel és Michalis Averoff mutatta be, akik egy kisméretű rákfaj, a Parhyale hawaiensis egyedeit felhasználva funkció-nyeréses és -vesztéses vizsgálatok kombinációját végezték el. És ezekből jól látszott (lásd alábbi ábrán), hogy ha az Ubx-et olyan elülső szelvényekben fejeztetem ki, ahol egyébként nem lennének jelen, akkor mind a csápok (Ant), mind az állkapocslábak (Mx2 és Mx3) külalakjukban járóláb jellegeket vesznek fel.

Akkor a funkcióvesztéses vizsgálatokat RNS-csendesítés (RNAi) kíséreltekkel végezték, de ezeknek van pár metodológiai problémája (például csak a legritkább esetben viszik le valóban nullára az expresszió szintjét), így a most szélsebesen terjedő CRISPR-alapú genomeditálással Patel-ék újravégezték az akkori kísérleteket. És ha már nekivágtak, akkor gyorsan odacsapták a többi Hox gént is, ezzel az utóbbi évek egyik legizgalmasabb Hox-témájú cikkéhez jutottak.
Az Ubx CRISPR-es kiütése még erősebben adta vissza a korábbi RNAi fenotípust, és azok az elülső járólábak, amelyek már az Ubx-t expresszáló szelvényekből nőttek ki, állkapocslábbá változtak. Az ún. anterior Hox gének (Deformed – Dfd, Sex combs reduced – Scr, Antennapedia – Ant) közül a Dfd és Scr hiányában az érintett szelvények nyúlványai csáp-szerűek lettek, illetve utóbbi esetében az egyik állkapocsláb pár járóláb-jellegű lesz.
Az Antp nélkül is hasonló transzformációkat látunk, például a második és harmadik torszelvény lábai poszteriorabb jelleget öltöttek.
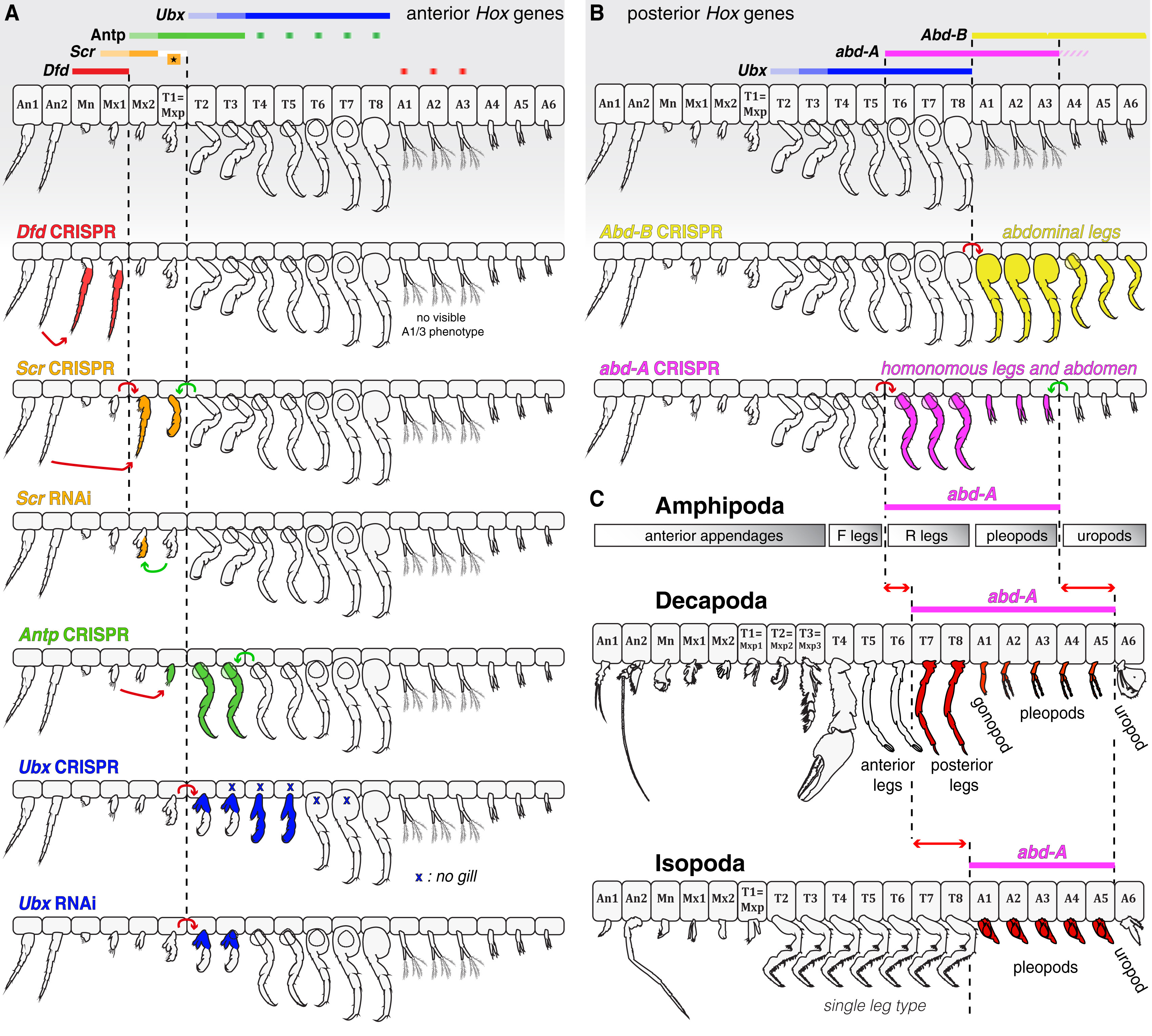
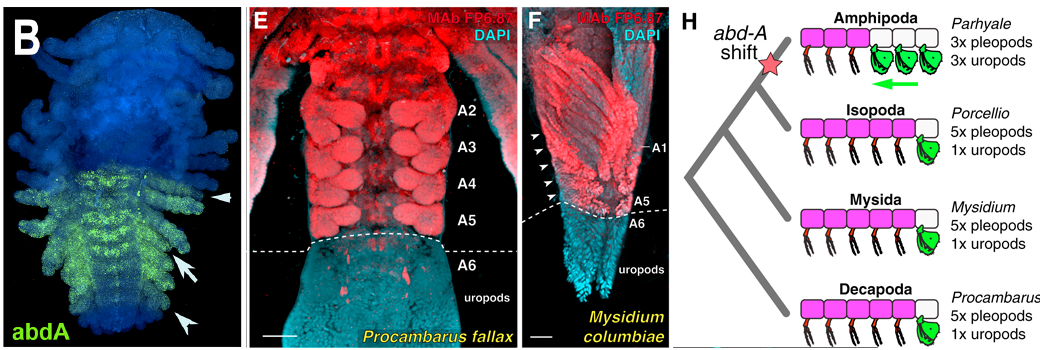
Az Ubx melletti két másik hátsó Hox-gén, az Abdominal-A (abd-A) és Abdominal-B (Abd-B) elvesztése pedig gyakran pont ellenkező hatást váltott ki, azaz anteriorizációhoz vezetett: például az Abd-B kiütésekor a potroh szelvényein az azokra jellemző úszólábak helyett ott is járólábak jelentek meg. Az abd-A hiánya kicsit komplexebb következményekkel járt, de ezek talán érdekesebbek, mert úgy tűnik, hogy evolúciósan is fontos dologról árulkodnak.
Először is, nagyon érdekes, hogy a hátsóbb torszelvények lábai az első torszelvények lábaihoz váltak hasonlóvá (ez elmegy anteriorizációnak). Viszont a potroh elülső szelvényeinek úszólábai olyan ún. uropodiumokká váltak, amelyek korábban csak az utolsó három szelvényen voltak megfigyelhetők (a lemezszerűen kiszélesedett uropodiumok a farok evező és kormányzószerkezetét alkotják). Ez pedig már egy olyan homeotikus mutáció, aminek nagyon is van evolúciós vonatkozása.
Az uropodiumok száma ugyanis rendszertani csoportonként változik, és míg számos rákfajban csak az utolsó potroh-szelvényen van uropodium, az Amphipoda rend esetében (ahova hősünk, a P. hawaiensis is tartozik) az utolsó három szelvény ilyen. Ráadásul, míg a többi csoportban az abd-A expressziója az utolsó potroh-szelvény (A6) kivételével a teljes potrohban megfigyelhető (alábbi képen pirossal látjuk ezt a Decapoda és a Mysida rend egy-egy képviselője esetén), az Amphipodáknál az A5 és A4 szelvényekben sokkal gyengébb abdA szintet látunk, mint az anteriorabb szelvényekben. Vagyis, nagyon abba az irányba mutatnak az adatok, hogy az Amphipodák uropodiumjai egy klasszikus, de evolúciósan konzervált, homeotikus mutáció eredményeinek tűnnek.
Egy izgalmas lappal bővült tehát a Hox-sztori, ami jól kiegészíti például a korábbi évek ugróvillás és vízipoloskás történeteit. És persze a puszta identitás-meghatározás nem az egyetlen dolog, amit a Hox gének végeznek: mint azt a tücskök, vagy méhek esetében látható, hogy az egyes végtagok hogyan nyerik el végső alakjukat, az egyes ízek hogyan fejlődnek, az is ezektől a különleges génektől függ. Már csak azt kell megértenünk, hogy pontosan hogyan és milyen géneket befolyásolnak ők maguk, mert a hosszú évtizedek óta tartó vizsgálódás ellenére, meglepően keveset tudunk erről.
Liubicich DM, Serano JM, Pavlopoulos A, Kontarakis Z, Protas ME, et al. (2009) Knockdown of Parhyale Ultrabithorax recapitulates evolutionary changes in crustacean appendage morphology. PNAS 106(33): 13892-6.
Pavlopoulos A, Kontarakis Z, Liubicich DM, et al. Probing the evolution of appendage specialization by Hox gene misexpression in an emerging model crustacean. PNAS 106(33): 13897-13902.
Martin A, Serano JM, Jarvis E, Bruce HS, Wang J, et al. (2015) CRISPR/Cas9 Mutagenesis Reveals Versatile Roles of Hox Genes in Crustacean Limb Specification and Evolution. Curr Bio doi:10.1016/j.cub.2015.11.021.
Valami gond van a régebbi cikkek linkjeivel.
Átlépek pl a „Méhláb és Hox gének” cikkre, de onnét már nem lehet továbblépni a „méhcsaládösszeomlás enigmájára”.
@unpopular.puffin: Ez sajnos a pár évvel ezelőtti blog-költöztetés hatása – a belső linkek teljesen összeomlottak akkor és hát esélyünk is, az összeset visszaírni :-///. De a poszt meg van, ha a belső keresőben nézed az összeomlást”, vagy Google-ben „összeomlás site:criticalbiomass.blog.hu” parancssorral, akkor ki kell jöjjön azért.
Akiket érdekel, az új Developmentben pont van egy Nipam Patel interjú, erről a munkáról is: dev.biologists.org/content/142/24/4189.full