
 A szárazföld meghódítása nemcsak az úszás helyett mászáshoz alkalmazkodott végtagokat igényelt, de a korábbi szaporodási stratégiák radikális újragondolását is szükségessé tette a gerincesekben.
A szárazföld meghódítása nemcsak az úszás helyett mászáshoz alkalmazkodott végtagokat igényelt, de a korábbi szaporodási stratégiák radikális újragondolását is szükségessé tette a gerincesekben.
Hiszen míg a vizes közegben az ivarsejtek szabadon engedésével le lehetett tudni az evolúciós sikerhez szükséges hajcihőt, addig a szárazföldön valami közvetlenebb módon kellett célba juttatni őket. És ez a közvetlenebb mód lett a külső ivarszervek kialakulása, amely hímek esetében a péniszt jelenti.
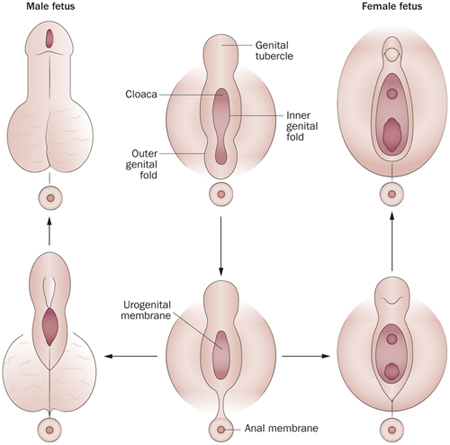 Ha egy standard emlőst, például egy embert veszünk alapul, akkor jól követhető a külső ivarszervek fejlődésében is, hogy a kezdeti embrió „bipotenciális” alapokon dolgozik, vagyis olyan ivarszerv-kezdeményeket hoz létre, amelyekből mind férfi, mind női megfelelők létrejöhetnek és csak a fejlődés későbbi szakaszában, az ivari kormoszómák által szabályozott hormonális hatásokra dől el, hogy pontosan merre fut a fejlődési program.
Ha egy standard emlőst, például egy embert veszünk alapul, akkor jól követhető a külső ivarszervek fejlődésében is, hogy a kezdeti embrió „bipotenciális” alapokon dolgozik, vagyis olyan ivarszerv-kezdeményeket hoz létre, amelyekből mind férfi, mind női megfelelők létrejöhetnek és csak a fejlődés későbbi szakaszában, az ivari kormoszómák által szabályozott hormonális hatásokra dől el, hogy pontosan merre fut a fejlődési program.
A leendő külső ivarszerv először az ivargumó (genitle tubercle – GT) formájában jelenik meg, amely közvetlenül az embrionális kloáka fölött jön létre (utóbbi, a legtöbb emlősben később egy válaszfal révén leválik a végbélnyílásra). A megfelelő fejlődési program révén, aztán a mellékelt ábrán jól követhető módon kialakulnak a külső ivarszervek, a GT pedig ennek az ivarszervnek a legérzékenyebb régiójává alakul.
Történetünk szempontjából azonban mindez, ha nem is lényegtelen, de másodlagos, mert a polihisztor Cliff Tabin csoportja most annak ment utána, hogy különböző gerinces csoportokban honnan ered a GT és milyen hatásokra alakul egyáltalán ki.
A sztori külön csavarja, hogy számos hüllő-faj nem egy, hanem két darab külső ivarszervvel rendelkezik (a kétkedők itt megtekinthetik a dolgot), szakzsargonban ezeket hemipénisznek nevezzük. (A részletek iránt érdeklődők érdekében: párzáskor csak az egyiket használják.)
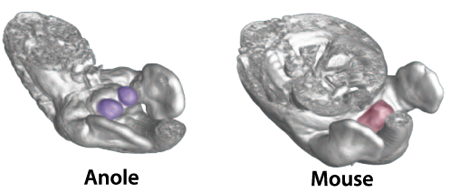
Ha nagyfelbontású CT képeken követjük az ivarszervek kialakulását, akkor jól látható, hogy míg az egerek esetében a GT a farokbimbó alatt jelenik meg, addig hüllőkben a hemipénisz (hp) kezdeménye a két hátulsó végtagbimbó tövében – a csökevényes hátsó végtaggal rendelkező pitonoknál az ivargumó és a végtagbimbó kb. ugyanakkora, míg a teljesen hátsó végtag nélküli fajoknál kvázi a végtagbimbó helyén jelenik meg az ivargumó.
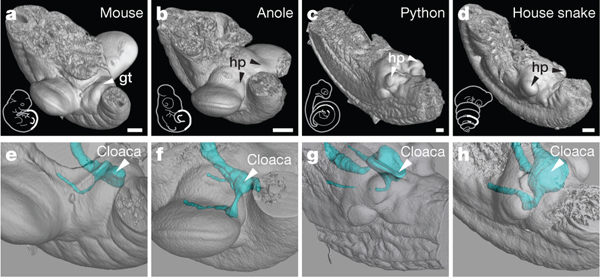
Annak eldöntésére, hogy valóban közös eredetű sejtek hozzák tehát létre az emlős péniszt és farkat, illetve a hüllő hemipéniszt és hátsóvégtagot, a kutatók egyszerűen, zöld fluoreszcens fehérjével megjelölték azokat a régiókat az embrióban, ahol a farokbimbó, illetve végtagbimbó megjelennek.
Az eredmény itt is szemléletes lett: míg egérben csak a farokbimbó megjelölése esetében jelentek meg sejtek az ivargumóban, a végtagbimbó jelölésnél pedig nem, addig a vizsgált gyíkfajban épp fordítva történt.
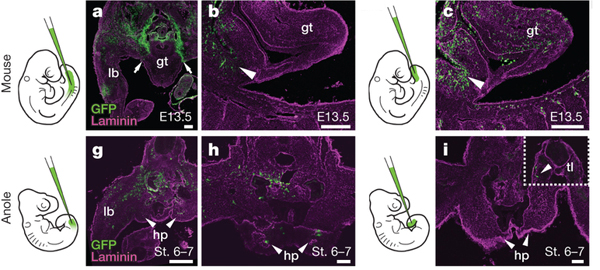
Mindez persze csak egy része a bizonyítékoknak: amikor megnézték, milyen gének fejeződnek ki egyes fajokban a végtagbimbóban, ivargumóban és farokbimbóban, szintén arra jutottak, hogy a hemipéniszek korai kezdeményeit alkotó sejtek leginkább a végtagbimbó sejtjeihez hasonlítanak (hogy aztán persze később nagyon eltérő identitást vegyenek fel).
Na de, mi indukálja vajon az egyes fajok különböző szöveteiben a GT kialakulását? Van-e valami univerzális szabály erre az indukcióra, vagy fajonként eltér a kiváltó ok?
Az egyik készenfekvő válasz az az embrionális kloáka lenne, hiszen ez mindig a GT/hp mellett található és ennek megfelelően kicsit hátrább is van egy egérben, mint a hüllőkben. Mi történik tehát, ha egy kloákát olyan helyre ültetünk át, ahol korábban nem volt? Mivel az emlős- és gyík-fajok nem praktikusak ilyenfajta kísérletekre, ezt csirkeembriók segítségével vizsgálták meg (csirkékben, egyébként, a kloáka köztes helyen van és kis mértékben mind a végtag-, mind a farokbimbó részt vesz az ivargumó kialakításában). Ha a kísérleti alanyokban a hátsó végtagbimbóra, vagy a farokbimbó egy távoli részére ültették a kloáka-szövetet, lokális, ivargumó szerű képződmények jelentek meg, amelyek ráadásul olyan markereket is fejeztek ki, mint a leendő ivarszervek.
Jól látszik tehát, hogy ahogy az evolúció során az „ivargumó-szervező központ”, vagyis az embrionális kloáka helyzete eltolódott a fejlődő magzatban, a külső ivarszervek sejtjeinek forrása is megváltozott, bár kialakulásuk, indukciójuk logikája ugyanaz maradt.
Természetesen a fejlődés későbbi szakaszaiban is jó alanya maradt az ivarszerv az evolúciós változásoknak, de az egy másik történet, amiről már emlősök esetében itt és itt, a madarakéban pedig itt írtam.
Tschopp P, Sherratt E, Sanger TJ, Groner AC, Aspiras AC, et al. (2014) A relative shift in cloacal location repositions external genitalia in amniote evolution. Nature doi: 10.1038/nature13819.
Elveszítettem a fonalat. Akkor most végtagbimbókból alakult ki az ivarszerv, vagy kloákából?
@Gazz: A kloáka indukálja a környező szöveteket (amelyek, feltételezem, valamennyire kompetensek kellenek legyenek, hogy ezt az indukciót érzékeljék), és azok sejtjei hozzák létre az ivargumót. Ha a közeli kompetens szövet a végtagbimbó, akkor annak a sejtjei, ha pedig a farokbimbó, akkor annak a sejtjei.
„radikális újragondolását is szükségessé tette”
Ez jól hangzó irodalmi megfogalmazás, de szerintem a laikusokban az ilyen mondatok adnak okot a félreértésre. Ugyanis ki gondolta újra? Az evolúció? Az evolúció nem gondolkodik se előre se hátra.
@G. M. E.: OK, ez jogos. Legközelebb duplán figyelek :-).