
 Az emberré válás egyik kulcskérdése a modern emberi agy kialakulása, ennek megfelelően ahogy egyre több genomi adat kezd a rendelkezésünkre állni, úgy kezdik el egyre többen nagy lelkesedéssel olyan gének/szabályozó elemek keresését, amelyek megmagyarázhatják az emberi agy méretének impresszív evolúciós növekedését.
Az emberré válás egyik kulcskérdése a modern emberi agy kialakulása, ennek megfelelően ahogy egyre több genomi adat kezd a rendelkezésünkre állni, úgy kezdik el egyre többen nagy lelkesedéssel olyan gének/szabályozó elemek keresését, amelyek megmagyarázhatják az emberi agy méretének impresszív evolúciós növekedését.
Ezeknek a kutatásoknak az eredményeit ismertető tudomány-rovatos újságcikkek általában előszeretettel szeretnek bombasztikus címekkel operálni és könnyedén dobálóznak az olyan nagyívű megállapításokkal, hogy „megtalálták az emberi agy komplexitásáért” felelős gént. A valóság persze az, hogy ezernyi ilyen apró „kulcsmutáció” lehet a genomban és ezek együttes hatása, ami az agy komplexitásához vezet – ráadásul lehet, hogy bizonyos mutációk csak adott kombinációban vizsgálhatók, önmagukban nem, ami tovább bonyolítja a kísérletek tervezését illetve értelmezését.
Az utóbbi hetekben két, az emberi agy evolúciójában szerepet játszó mutációkkal foglalkozó cikk is nagyobb figyelmet kapott, ezeket veséznénk ki sebtiben. Ami közös bennük, hogy humán-specifikus DNS szakaszokat (egyik esetben egy szabályozóelemet, a másik esetben egy teljes gént) fejeztek ki egér agyban és ennek eredményeként fellépő változásokat jellemezték. Ez a megközelítés nem új, korábban sikeresen alkalmazták a „beszédgénként” sztárolt FOXP2 esetében is. A kísérleteknek az a praktikus logikája van, hogy az egér viszonylag egyszerű agykérgében könnyen kimutatható, ha valami statisztikailag szignifikáns változás következik be egy humán szekvenciák hatására, plusz – nem elhanyagolható módon – kevesebb etikai problémával kell szembesülni, mint ha főemlősökön kísérleteznének. (Lásd még az asztrocita-kimérák esetét is.)
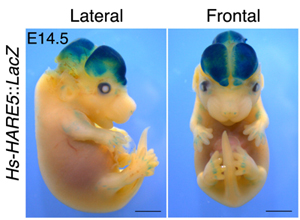 Ha kronológiailag megyünk, a korábbi cikk a Current Biology-ban jelent meg és egy olyan konzervált, de nem kódoló DNS-szakasz funkcióját boncollgatja, ami a csimpánz-ember evolúciós vonal szétválása után a mi oldalunkon feltűnően sok mutációt szedett össze.
Ha kronológiailag megyünk, a korábbi cikk a Current Biology-ban jelent meg és egy olyan konzervált, de nem kódoló DNS-szakasz funkcióját boncollgatja, ami a csimpánz-ember evolúciós vonal szétválása után a mi oldalunkon feltűnően sok mutációt szedett össze.
A Hs-HARE5, ha egy egyszerű riporter-génhez kötjük (pl. a kék festést lehetővé tevő β-galaktozidázt kódoló LacZ) jellegzetesen az agyban mutat expressziót.
Helyzete alapján feltételezni lehetett, hogy a Wnt-jelátvitelben játszik valami szerepet, ugyanis a Frizzled-8 (Fzd8) nevű génhez relatíve közel található és ez utóbbi gén a Wnt-jelátvitel központi receptora.
A közvetett közelségen túl ráadásul a kromatin szerkezetét vizsgáló eljárások is alátámasztották, hogy a kromatin aktív működése során, a HARE5 és FZD8 egymással közvetlen kapcsolatba kerülnek, azaz előbbi pont úgy viselkedik, mint az utóbbi egyik szabályozóeleme.
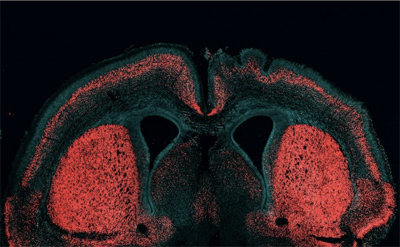
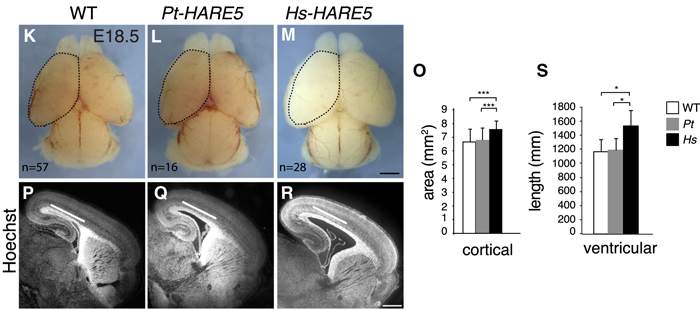
A kérdés persze az, hogy mi történik, ha mondjuk a humán HARE5 működését a csimpánz ortológ (Pt-HARE5) működésével vetjük össze? Ennek megválaszolására létrehoztak olyan transzgénikus egereket, ahol vagy a csimpánz, vagy a humán szekvencia hajtotta meg az egér saját Fzd8 génjét. Az eredmény: a Pt-HARE5 esetében az egerek agya lényegében pont olyan, mint a kezeletlen kontrollokéban. Ezzel szemben a Hs-HARE5 szekvenciával rendelkezők agymérete megnőtt – ha nem is valami fantasztikusan, elképesztően nagyon, annyira biztosan, hogy az szignifikáns legyen.
(Ez egyébként szinte biztosan nem az utolsó ilyen típusú cikk: pár hónapja egy konferencián David Kingsley nagyon hasonló logikával egy másik humán specifikus DNS-szakasz elemzését mutatta be (hasonló eredménnyel), csak az még nem lett azóta sem publikálva.)
A másik, a Science hasábjain bemutatott kísérlet során először rendszertanilag távolabbi kategóriákat vetettek össze: történetesen magát az egeret, az emberrel. Viszont ezúttal nem egyszerű genomi különbségekre voltak kíváncsiak, hanem azt nézték, hogy azokban a sejtekben, amelyek a fejlődés során a neuronokak hozzák létre (ún. radiális gliasejtek), milyen génexpressziós különbségek fedezhetők fel egér és ember között.
 Ezen belül azonban leginkább azokra a génekre koncentráltak, amelyek csak az emberben vannak jelen, más fajokban pedig nem. Így jutottak a nem túl költői hangzású ARHGAP11B génhez, amely a sejten belüli szignalizációban központi szerepet játszó G-fehérjék egyik szabályozóját kódolja.
Ezen belül azonban leginkább azokra a génekre koncentráltak, amelyek csak az emberben vannak jelen, más fajokban pedig nem. Így jutottak a nem túl költői hangzású ARHGAP11B génhez, amely a sejten belüli szignalizációban központi szerepet játszó G-fehérjék egyik szabályozóját kódolja.
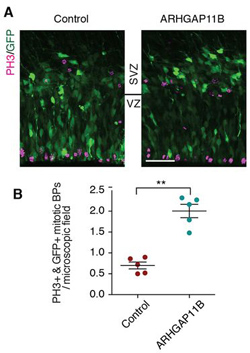
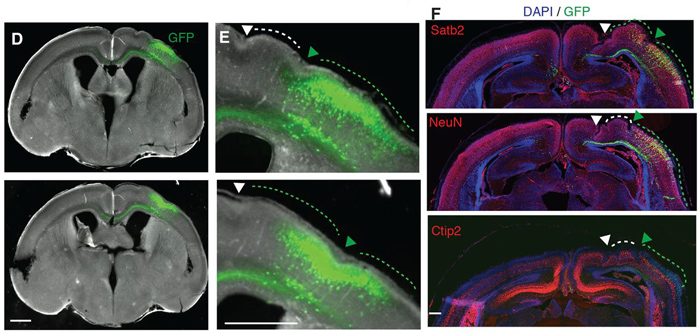
Ezt az emberi gént aztán kifejeztették az egér radiális-glia sejtjeiben, amelynek az egyik folyománya, hogy azok osztódási aktivitása jelentősen megnövekedett. Ennek pedig részleges eredménye és következménye az lett, hogy ha fejlődő egerek agyának csak egy bizonyos területére elektroporálták (fluoreszcens GFP-vel együtt) az ARHGAP11B gént, akkor az a terület morfológiailag is elkezdett különbözni a környező (kvázi kontrollként működő) régióktól.
Bemélyedések jelentek meg rajta, amelyek – ha hiszünk a szerzőknek – már már kvázi barázdáknak felelnek meg. (A barázdáltság azért fontos jellemzője az emberi agynak, mert ezek révén egységnyi térfogatba sokkal nagyobb agykéreg felületet lehet befértetni.)
Vagyis az ARHGAP11B megjelenése szintén kritikus eleme lehetett annak a kirakósnak, ami végül a modern Homo sapiens agyának kialakuláshoz vezetett.
Boyd JL, Skove SL, Rouanet JP, Pilaz LJ, Bepler T, at al. (2015) Human-Chimpanzee Differences in a FZD8 Enhancer Alter Cell-Cycle Dynamics in the Developing Neocortex. Curr Biol pii: S0960-9822(15)00073-1.
Florio M, Albert M, Taverna E, Namba T, Brandl H,et al. (2015) Human-specific gene ARHGAP11B promotes basal progenitor amplification and neocortex expansion. Science pii: aaa1975.