
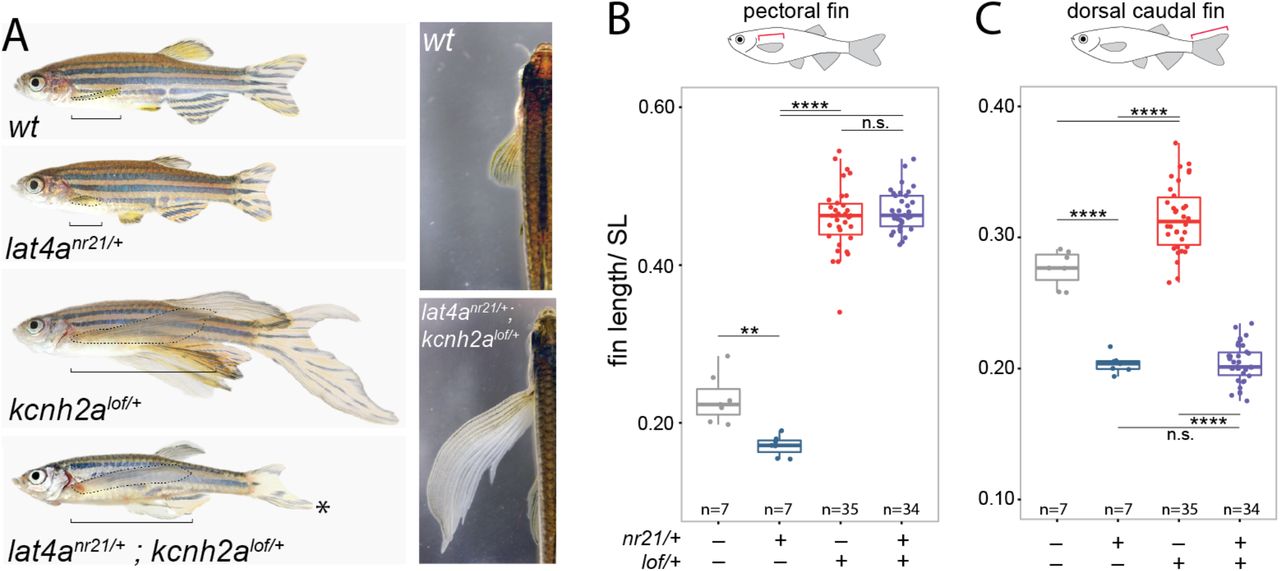
A homoszexualitás napjainkban egy megosztó téma. Országunkban jelenleg különösen aktuális kérdés: Elfogadható-e az azonos neműekkel folytatott intim kapcsolat, illetve szabad-e ennek létezéséről tudniuk a kiskorúaknak? Úgy tűnik, mostanság egyik sem kívánatos, köszönhetően egy új törvénymódosításnak. Noha ezt talán senkinek nem is kell részleteznem, a lényeget kiemelném.
…tilos tizennyolc éven aluliak számára pornográf, valamint olyan tartalmat elérhetővé tenni, amely a szexualitást öncélúan ábrázolja, illetve a születési nemnek megfelelő önazonosságtól való eltérést, a nem megváltoztatását, valamint a homoszexualitást népszerűsíti, jeleníti meg…
Javaslat módosítási szándék megfogalmazásához a T/16365. számú törvényjavaslathoz
Homoszexualitás és az emberi természet
A módosító hátterében minden bizonnyal az régóta hangoztatott érv áll, miszerint egy homoszexuális kapcsolat természetellenes (peccatum contra naturam, latin: természet ellen elkövetett bün), s ilyenektől védeni kell a fiatal generáció elméjét. De valóban természetellenes-e az azonos neműekkel folytatott viszony? Biológusként kicsit kötelességemnek érzem, hogy e témát pár szóban és pár példával körüljárjam.
Nem fogok pontos definíciókkal élni. A következőkben a “homoszexualitás” alatt egyfajta “nem heteroszexuális” viselkedést fogok érteni. Kezdjük ott, hogy magát a homoszexualitást is nehéz kategorizálni. Manapság ez egyfajta kizárólagosságot hordoz magában. A témával való kutatások többsége azonban inkább egy skála (pl. Kinsey-skála) mentén szokta osztályozni a szexualitást attól függően, hogy valaki kizálólagosan preferál egy nemet, netán csak fantázia szintjén, esetleg alkalmilag érdeklődik az egyik vagy a másik iránt (1. ábra).
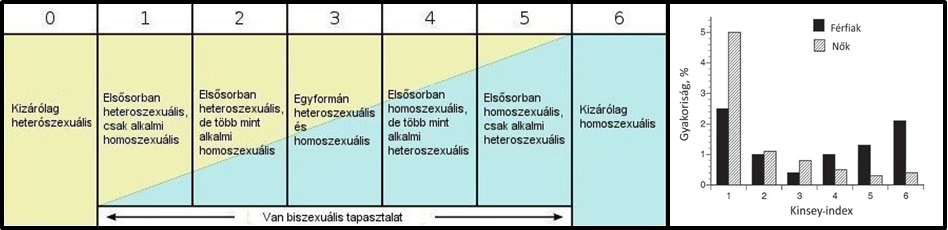
nyomán, forrás: Scheuring, 2014)
Például az ókori Görögországban és Rómában teljesen elfogadott volt házas férfiként, családapaként fiatal férfiú szeretőket tartani (bár Platón idejében indultak viták ennek “rendbontó” hatásairól). Más kultúrákban is komoly történelme van a homoszexualitásnak. Kínában már i.e. 600 körülről vannak feljegyzések homoszexuális viszonyokról, és ismeretesek egyéb, Közel- és Távol-Keleti példák is, mely nagyon fluid szexuális preferenciákról (vagy épp nemi szerepekről) számolnak be. E viszonyok morális megkérdőjelezése csak 1000-1200 évvel később, a keresztény és iszlám betörésével indul meg.
Szintén érdekes óceániai kultúrák koncepciója a nemi identitásról és szexuális preferenciáról. A polinéz nyelvekben nagyon régóta létezik szó “azonos neműek közötti intim kapcsolat”-ra (maori: takatāpui; hawaii: moe aikāne). James Cook legénységének beszámolói alapján ez teljesen elfogadott és stigma nélküli együttlét volt. Szintén érdekesség, hogy e népcsoportokban gyakran létezik a társadalmilag elfogadott, néha fontos kulturális szerepet betöltő “harmadik nem” megkülönböztetése (Hawaii: māhū; Cook szk.: ‘akava’ine; Tonga: fakaleitī; Tuvalu: pinapinaaine; Niue: fakafifine; Szamoa: faʻafafine). Ők férfiként saját nemükhöz vonzódnak, és számos tekintetben nőként viselkednek (egy széles spektrumban). Talán a transzneműség a legkifejezőbb szó, de nehéz lenne ezt teljes mértékben lefordítani a 21. századi LGBT paradigmára, és teljesen más kulturális identitás jellemzi például Szamoán a fa’afafinéket, akikről később még említést teszünk.
Tehát régóta léteznek kultúrák, ahol egyáltalán nem tabu az azonos neműek közötti kapcsolat. Vajon e népek elhajlása csak egy téves berögződés, netán valami sokkal általánosabb trend, mely túlmutat az emberi viselkedésen?
Homoszexualitás az állatvilágban?
Az utóbbi 20 év megfigyelései alapján ma már több mint 1500 faj esetében írtak már le homoszexuális viselkedést az állatvilágban (same-sex sexual activity: az egyed egy másik azonos nemű egyeddel létesít szexuális jellegű kapcsolatot). Akadnak köztük szép számmal rovarok, halak, kétéltűek, hüllők, madarak és emlősök is.
A házi juh (Ovis aries) esetében például a kosok egytizede elutasító nőstényekkel szemben, míg más kosokkal előszeretettel kezdenek ki. A Laysan albatrosz (Phoebastria immutabilis) Oahu szigetén fészkelő populációjánál a fészkelő párok kb. egyharmada nőstény-nőstény pár. Egyes megfigyelések alapján a zsiráfbikák is gyakrabban udvarolnak egymásnak mint nőstényeknek. És a főemlősök sem képeznek kivételt: japán makákók, gorillák és orángutánok esetében is leírtak már azonos nemű közösüléseket.
Az ember egyik legközelebbi rokona, a törpecsimpánzok (Pan paniscus) példája maga a szodomizmus netovábbja. Nemcsak, hogy nagyon is “öncélúan” élnek a szexuális aktivitással, hanem itt a biszexualitás a norma, az aktusok több mint fele kettő vagy több nőstény között történik. De a hímek közötti szex is bevett szokás! Társadalmukban a szex sokkalta több szereppel bír mint az utódnemzés funkciója. De erre majd később visszatérünk.

És ez csak pár kiragadott példa volt. Tehát közel sem állítható az, hogy a homoszexualitás pusztán valami “mentális perverzió” lenne. Ez egy viszonylag gyakori, sőt mondjuk ki, ez igenis természetes és gyakran előforduló viselkedési forma az állatvilágban is.
Isten és Darwin nevében
Akkor miért tarthatjuk természetellenesnek e viselkedést? Ha amúgy az állatvilágban gyakori stratégia, akkor lehet hogy pont a kulturális hátterünk miatt? Nem szeretnék valakinek a hitvilágába belegázolni s azt a végletekig boncolni, de azért spekulálhatunk (ilyenek ezek a spekulánsok!), mint egy evolúcióbiológus. Innen kapargatva a felszínt talán jobban érthető lesz, hogy miért tapasztalható meg egyfajta értetlenség és mélyen gyökerező ellenszenv a homoszexualitással szemben.
Valószínűsithető, hogy a homoszexualitás morális megkérdőjelezése főképp az ábrahámi vallásoktól (pl.: zsidó, keresztény, iszlám) és azok későbbi befolyásából (nyugatiosodásból, kolonizáció) ered. Ironikus módon ugyanis e világvallások értékrendje meglepően “darwinista”, legalábbis tartogat olyan alapvetéseket mely egy evolúcióbiológus fantáziáját is megmozgatja.
E vallások egyfajta adaptív funkciója, hogy egy összetartóbb és erősebb humán csoport megszervezésére törekszik egy “kulturális kódex” segítségével. Az egyik ilyen érdekes aspektus például a Nagy Testvér személye (nevezzük bármilyen néven is): az emberek többsége sokkalta jobban kooperál (vagy mellőzi a csalásokat bizonyos szituációkban) ha van egy külső megfigyelő, hisz az maga után vonhat egy későbbi megtorlást. Márpedig egy omnipotens entitás mindent lát és minden kihágást büntethet.
Egy másik ilyen, és mostani témánk szempontjából fontosabb, dogma a szex társadalmi funkciójának tisztázása a csoport érdekében. Ha különböző csoportok versengenek, akkor fontos, hogy melyik csoport szaporodik gyorsabban. Az lesz előnyben a versenytársakkal szemben. A feltételezés esetünkben pedig az, hogy nagyobb szaporodási rátát lehet elérni azzal, ha mindenféle “öncélú szexuális tevékenység” helyett a ráfordítható energia egy családba, illetve egy exkluzív partnerrel való utódgondozásba megy. Részben ezért is válik természetellenessé és elítéltté egy homoszexuális kapcsolat: nincs belőle utód, a csoport számára pedig ez nem feltétlen előnyös.
A darwini paradoxon és lehetséges megoldások
De nem csak társadalmi, hanem evolúciobiológiai szempontból is paradoxonnak tűnhet az azonos neműekkel folytatott viszony. Valószínűleg van a viselkedésnek egy öröklődő, genetikai háttere. Scheuring István előadása és korábbi Természet Világa cikke ezt sokkal élvezetesebben vesézi ki, csak ajánlani tudom (és nem tagadnám, hogy nagy hatással voltak e posztra is). Viszont ha ez örökletes, és homoszexuális embereknek kevesebb utódja születik (nem feltétlen nulla), akkor a “homoszexuális” génnek ki kellene szelektálódnia. De nem ezt látjuk. Ez pedig egy érdekes evolúciós kérdés: Miért nem tűnik el a homoszexuális viselkedés a populációkból?
Lehet, hogy a homoszexualitásnak van valamilyen közvetett előnye? A Laysan albatrosz esetében a kiváltó ok a hímek hiánya: a nőstény-nőstény párok költési sikere noha kicsi (az apa ilyenkor egy alkalmilag félrelépő hím), de a sikeres költés esetén több esélyük van a következő idényben hím párt fogni maguknak. Egyes, hímeket teljesen nélkülöző és önmaguk megtermékenyítésére képes (ún. partenogenetikus) gyíkfajoknál (pl. Aspidoscelis neomexicanus) a nőstények közötti szex valószínűleg elengedhetetlen az önmegtermékenyítés a beindításához.
Az ember esetében rokonszelekció gyanúja merül fel mint lehetséges mechanizmus. Ha például egy homoszexuális egyed utódnevelés helyett inkább a rokonait segíti, azzal továbbra is hozzájárulhat a hajlamosító gén terjedéséhez. Az ugyanis a rokonban is nagy eséllyel megtalálható. Pusztán arra van szükség, hogy a segítség mértéke és a rokonsági fok meghaladja a homoszexuális egyed költségét. Ez a közvetlen előny már fenntarthatja a homoszexualitást a “hordozók” segítésével.
Vannak eredmények mely ezt a rokonszelekciós narratívát erősíthetik, bár korántsem beszélhetünk átfogó bizonyítékokról. Több kutatás (például ez is) kimutatta, hogy heteroszexuálisokhoz képest a homoszexuálisok nőrokonainak magasabb az átlagos utódszáma; ez egy szaporodásbeli előny, és X-kromoszómához kötött öröklődésre is utal. Azt sajnos nem sikerült kideríteni, hogy ez a női rokonok felé nyújtott segítség pontosan miben jelenik meg.
Talán ebből a szempontból érdekesek a korábban már említett szamoai fa’afafine-k (ők a már említett harmadik nem tagjai, nőies férfiak akik férfiakhoz vonzódnak). Esetükben kimutathatő, hogy sokkal jobban aggódnak a testvéreikért mint a heteroszexuálisok. Döntési kényszerek esetén is jobban preferálják azon rokoni kategóriák támogatását, amely a genetikai rokonságot és a rokoni előnyt maximalizálja. Ezek a kategóriák éppenséggel a lánytestvérek, azok gyermekei, és ott is elsősorban a lányok. És hogy miért a nők? Az anya kiléte sosem kérdés, így rokonság mindig biztosabb lesz a női vonalon.
Ugyanakkor érdemes megemlíteni, hogy a fa’afafine-k esetében talált eredmények a rájuk jellemző nőies viselkedéssel és a társadalmi háttérrel is összefüggésbe hozhatók, nem feltétlen csak a homoszexualitással. Nyugati társadalmak és Japán homoszexuális férfiainál például nem figyelhetők meg ilyen döntési trendek. Tehát elképzelhetők más, genetikai és nem rokonszelekción alapuló mechanizmusok is amik fenntartják a homoszexualitást.
És ha már társadalom, szóba jönnek a törpecsimpánzok. Náluk a főképp alkalmi, homoszexuális kapcsolatoknak elsősorban feszültségcsökkentő és békítő szerepe van. Ez a társadalmi csillapító szerep hozzájárulhat a stabil közösségek megszervezéséhez. Tekintve, hogy az ember egyik legközelebbi rokonáról van szó, felmerül a kérdés: emberelődöknél, netán korábban emlegetett emberi kultúráknál ez mekkora szerepet játszhatott az azonos neműek közötti kapcsolat a nagyobb egyedcsoportok, társadalmak kialakításában?
Zárszó
A sokféleség Földünk és annak élővilágának velejárója. Minden fajon belül találunk kisebb és nagyobb egyedeket, színváltozatokat. A szexuális preferencia is csak egy ilyen dolog, nem egy természetellenes viselkedés. Érthető, hogy társadalmi közegünkben miért tűnik értelmetlennek, azonban ha jobban utánajárunk, rájöhetünk, hogy egyrészt a homoszexualitás nem ritka, másrészt akadnak logikus magyarázatok létezésére.
És itt szó sincs népszerűsítésről. Ez egy jelenség, a létezését tudomásul kell venni, pláne ha az továbbra is hátrányos megkülönböztetéshez vezet. Nem népszerűsíteni kell, nem is kell félni tőle, hanem megérteni. Ha valaki engem (mint evolúcióbiológust) kérdez, a fiatalabb generációt megakadályozni a világ és a sokféleség megértésében igenis hátrányos lehet. Pláne ha egy jelenségnek megvan a helye a nemcsak a természetben, de társadalmunkban is.
Scheuring, I. (2014) ‘A homoszexualitás evolúciógenetikai háttere’ Természet Világa, 145 (11). pp. 496-500.
Bailey, J. M., Dunne, M. P. and Martin, N. G. (2000) ‘Genetic and environmental influences on sexual orientation and its correlates in an Australian twin sample.’, Journal of Personality and Social Psychology, 78(3), pp. 524–536. doi: 10.1037/0022-3514.78.3.524.
Iemmola, F. and Camperio Ciani, A. (2009) ‘New Evidence of Genetic Factors Influencing Sexual Orientation in Men: Female Fecundity Increase in the Maternal Line’, Archives of Sexual Behavior, 38(3), pp. 393–399. doi: 10.1007/s10508-008-9381-6.
VanderLaan, D. P., Petterson, L. J. and Vasey, P. L. (2017) ‘Elevated Kin-Directed Altruism Emerges in Childhood and Is Linked to Feminine Gender Expression in Samoan Fa’afafine: A Retrospective Study’, Archives of Sexual Behavior, 46(1), pp. 95–108. doi: 10.1007/s10508-016-0884-2.
VanderLaan, D. P. and Vasey, P. L. (2014) ‘Evidence of Cognitive Biases for Maximizing Indirect Fitness in Samoan Fa’afafine’, Archives of Sexual Behavior, 43(5), pp. 1009–1022. doi: 10.1007/s10508-014-0288-0.
De Waal, F. B. M. (2006) ‘Bonobo Sex and Society’, Scientific American, 16(2), pp. 14–21. doi: 10.1038/scientificamerican0606-14sp.
Wilson, D. S. (2002) Darwin’s Cathedral. University of Chicago Press. doi: 10.7208/chicago/9780226901374.001.0001.
Young, L. C. and VanderWerf, E. A. (2014) ‘Adaptive value of same-sex pairing in Laysan albatross’, Proceedings of the Royal Society B: Biological Sciences, 281(1775), p. 20132473. doi: 10.1098/rspb.2013.2473.



 Ha Darwinról és hozzá közel álló madarakról beszélünk, akkor talán nem is feltétlenül a kötelező újggyakorlatként felemlegetett galapagosi pintyeket (aka. “Darwin pintyek”)
Ha Darwinról és hozzá közel álló madarakról beszélünk, akkor talán nem is feltétlenül a kötelező újggyakorlatként felemlegetett galapagosi pintyeket (aka. “Darwin pintyek”) 


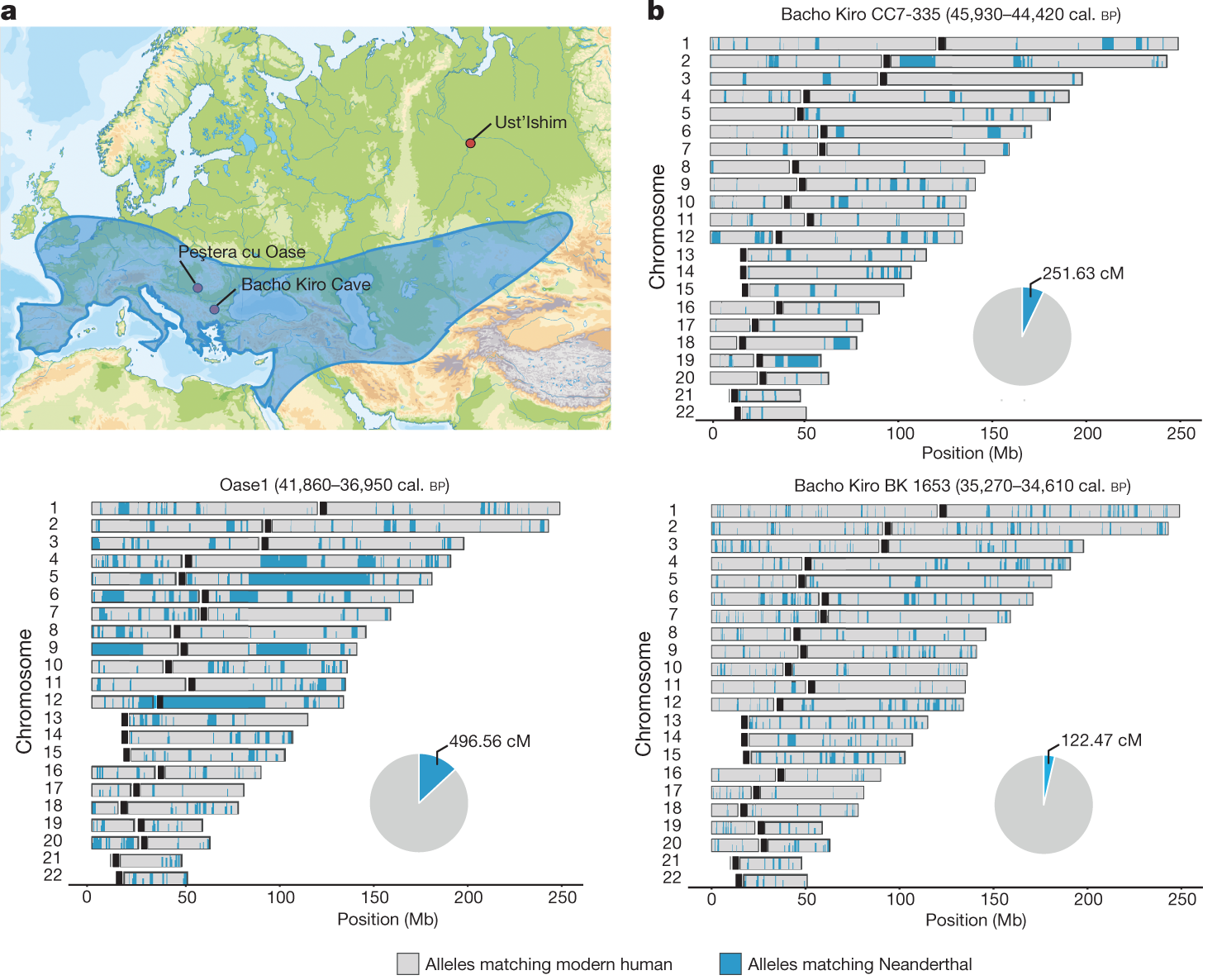
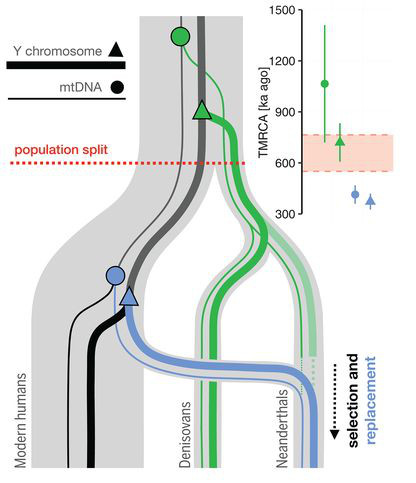
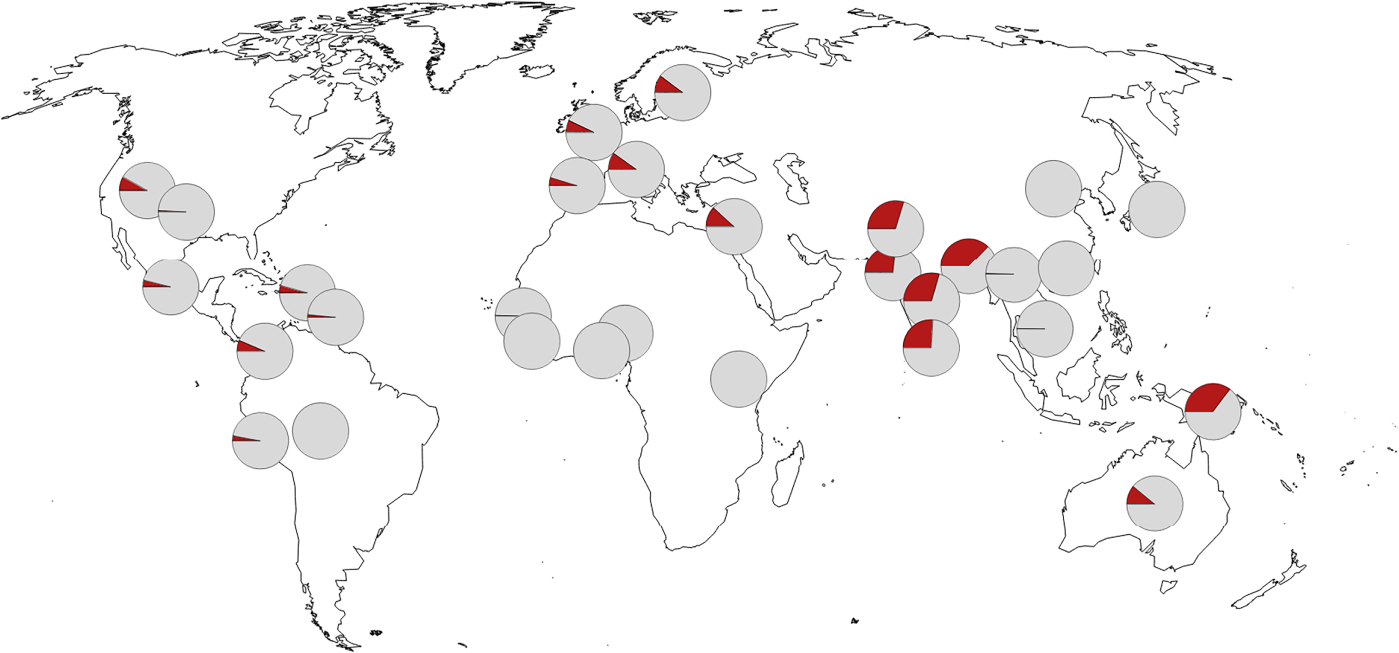

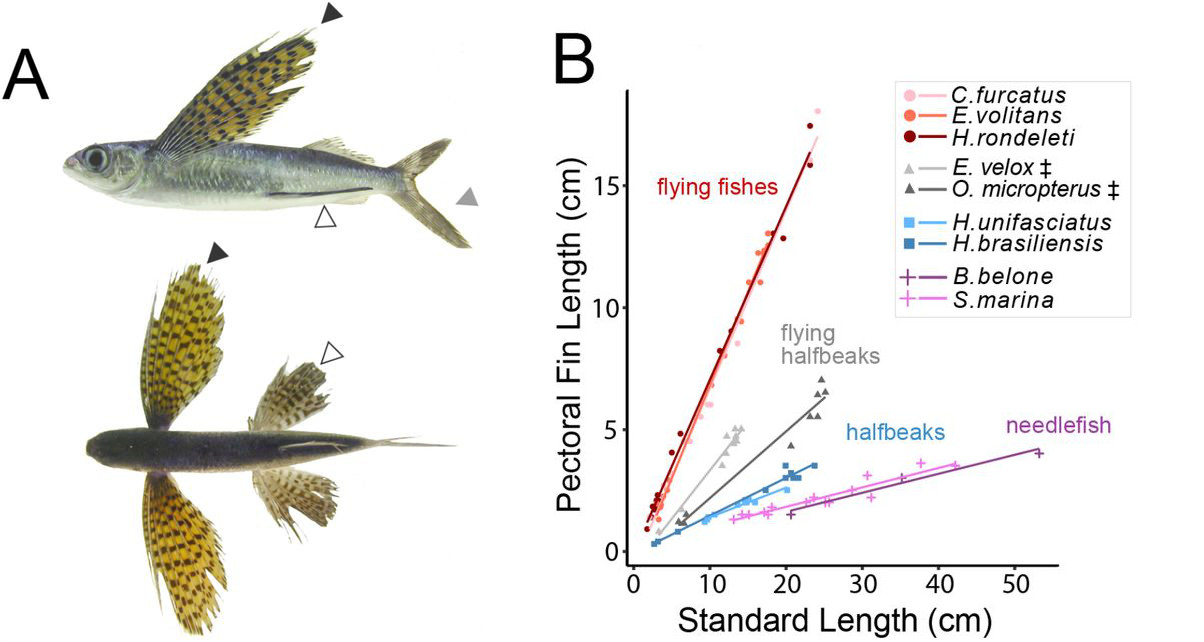

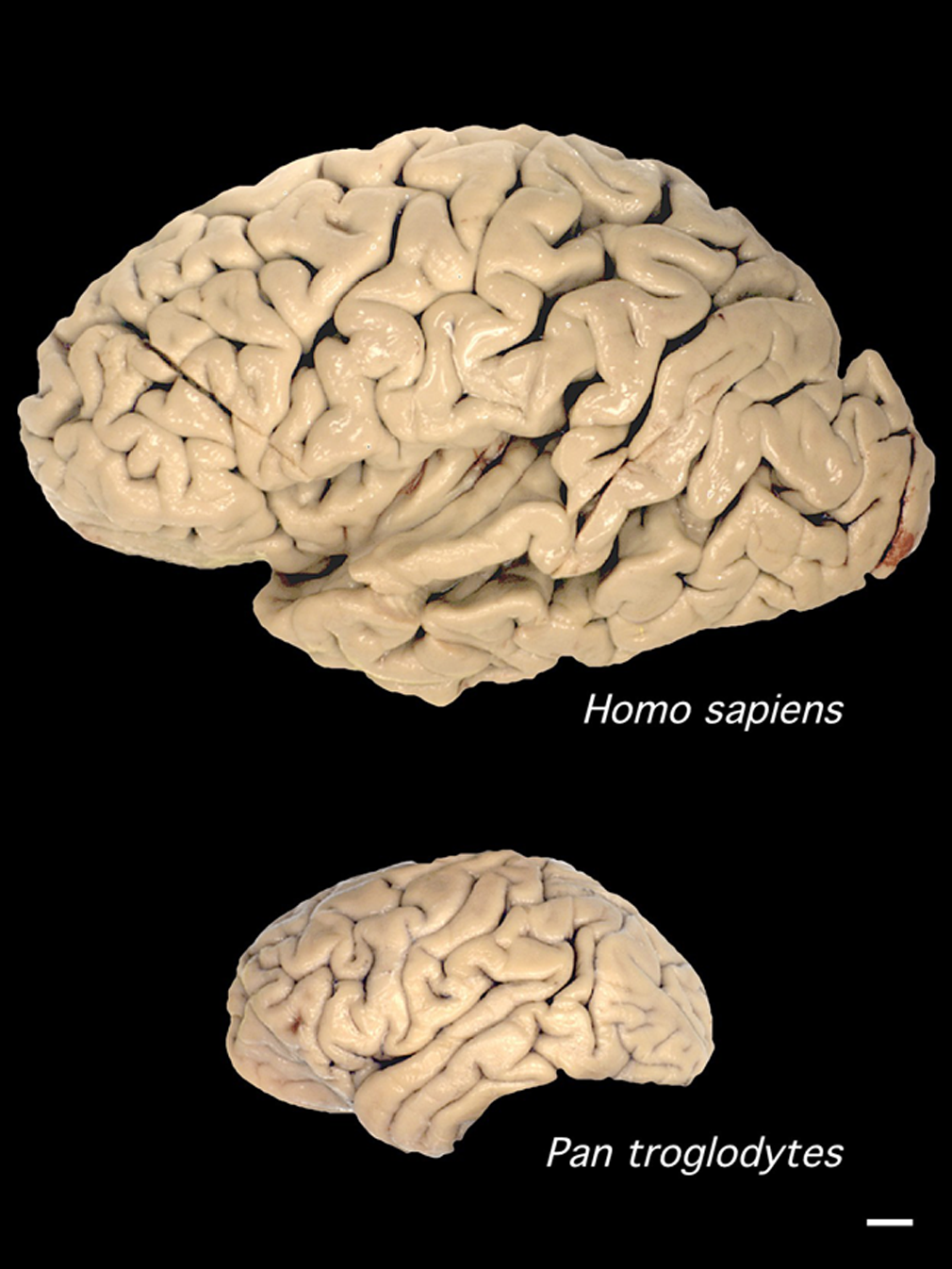 Bár a Fajok eredete igazából nem az ember eredetéről szól, mégis mindenki tudta és érezte már a megjelenésekor, hogy ez az az igazi kérdés, amit a benne leírt tudományos elmélet feszeget. Darwin, a rá jellemző visszafogottsággal, ebben a könyvében csak egy rövidke utalást tesz a kérdésre (“Fény derül majd az ember eredetére és történetére. (Light will be thrown on the origin of man and his history)”, amolyan beharangozóként későbbi, másik főművéhez, ami Az ember származása és az ivari kiválás lenne. Mindez jól tükrözi, hogy már bő másfél évszázada (ha pont nem “már a görögök óta”) az egyik legfontosabb kérdés, ami sok biológiai kutatás középpontjában is áll, arra keresi a választ, hogy mi az, ami minket fajként különlegessé tesz és megkülönböztet más fajoktól.
Bár a Fajok eredete igazából nem az ember eredetéről szól, mégis mindenki tudta és érezte már a megjelenésekor, hogy ez az az igazi kérdés, amit a benne leírt tudományos elmélet feszeget. Darwin, a rá jellemző visszafogottsággal, ebben a könyvében csak egy rövidke utalást tesz a kérdésre (“Fény derül majd az ember eredetére és történetére. (Light will be thrown on the origin of man and his history)”, amolyan beharangozóként későbbi, másik főművéhez, ami Az ember származása és az ivari kiválás lenne. Mindez jól tükrözi, hogy már bő másfél évszázada (ha pont nem “már a görögök óta”) az egyik legfontosabb kérdés, ami sok biológiai kutatás középpontjában is áll, arra keresi a választ, hogy mi az, ami minket fajként különlegessé tesz és megkülönböztet más fajoktól. 


{kind=link}