Az emberszabású majmok és az emberek esetében közismert az utánozási hajlandóság (sőt, a jelek szerint mi magunk méginkább hajlamosak vagyunk erre, mint a csimpánzok). Az utódok szüleik modulatainak, mozgás kombinációinak megismétlésével gyakran olyan hasznos képességeket tanulnak el, amelyek később segíthetik boldogulásukat a populáció szociális hálójában, vagy valamilyen más szempontból könnyítik meg az életüket. Sokak szerint ez az utánozó-hajlam az emberi kultúrák kialakulásának elengedhetetlen feltétele volt.
S bár a komplex viselkedések utánzása, valószínűleg valóban leginkább az ember (és néhány esetben az emberszabásúak) doméniuma, több jel mutat arra, hogy az utánzási kézség már a nem emberszabású majmokban is kódolva van.

A PLoS Biology-ban megjelent cikk pár napos makákók esetében mutat ki "majmolási" hajlandóságot, amely azonban két hetes korra megszűnik. Bár a dolog valóban újdonság, nem teljesen váratlan, hiszen, mint arra a PLoS kommentátora is utal, már korábban ismert volt, hogy a nem emberszabású majmok is rendelkeznek ún. "tükör-neuronokkal" (mirror neurons), olyan különleges sejtcsoportokkal, amelyek nemcsak akkor aktivizálódnak, ha az állat egy adott mozgást végez, hanem akkor is, ha egy társát nézi, amint az a szóbanforgó mozgást végzi (kis túlzással olyan mintha az állat elképzelné, hogy ő mozog). S bár szinte kétségtelen, hogy ezeknek a sejteknek fontos szerepük van az utánzási viselkedés során, pontosan még tisztázandó, hogy pontosan mi a funkciójuk.
Ferrari PF, Visalberghi E, Paukner A, Fogassi L, Ruggiero A, et al. (2006) Neonatal Imitation in Rhesus Macaques. PLoS Biol 4(9): e302.
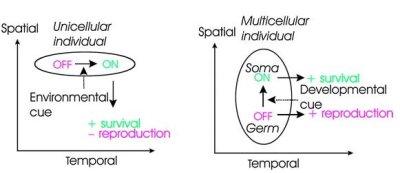
 Ritkán boncolgatott ténynek számít, hogy a nemek a társadalomban fele-fele arányban képviseltetik magukat, pedig érdemes feltenni a kérdést, hogy miért is van ez így? Most nem konkrétan arra gondolok, hogy az X és az Y kromoszómák szegregációja miért olyan amilyen (amely kétségtelenül megadja a mechanisztikus okát annak, hogy az emberek ill. általában az emlősök között miért ez a hímek és nőstények aránya), hanem hogy milyen evolúciós oka van annak, hogy az állatvilágban ez az arány a legelterjedtebb.
Ritkán boncolgatott ténynek számít, hogy a nemek a társadalomban fele-fele arányban képviseltetik magukat, pedig érdemes feltenni a kérdést, hogy miért is van ez így? Most nem konkrétan arra gondolok, hogy az X és az Y kromoszómák szegregációja miért olyan amilyen (amely kétségtelenül megadja a mechanisztikus okát annak, hogy az emberek ill. általában az emlősök között miért ez a hímek és nőstények aránya), hanem hogy milyen evolúciós oka van annak, hogy az állatvilágban ez az arány a legelterjedtebb.
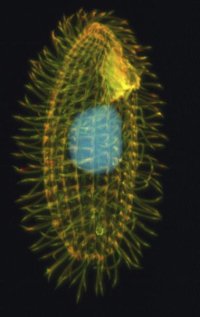 Minden élőlény genomja önmagában is izgalmas, hiszen bizonyos szempontból kódolja az illető faj egész történetét – bár kétségtelen, hogy ennek a történetnek a dekódolásához és megértéséhez sok más genomot is ismernünk kell. Az egysejtű eukarióták genetikai anyaga arról is mesél, hogy milyen lehetett az ősi eukarióták genomja, még a
Minden élőlény genomja önmagában is izgalmas, hiszen bizonyos szempontból kódolja az illető faj egész történetét – bár kétségtelen, hogy ennek a történetnek a dekódolásához és megértéséhez sok más genomot is ismernünk kell. Az egysejtű eukarióták genetikai anyaga arról is mesél, hogy milyen lehetett az ősi eukarióták genomja, még a  Állati szarvakról beszélve (már csak nyelvi okok miatt is) elsősorban mindenkinek a szarvas szokott beugrani, pedig az igazi nagymesterei a szarvnövesztésnek a bogarak között találhatók. A
Állati szarvakról beszélve (már csak nyelvi okok miatt is) elsősorban mindenkinek a szarvas szokott beugrani, pedig az igazi nagymesterei a szarvnövesztésnek a bogarak között találhatók. A 
 … avagy, mint az Economist is
… avagy, mint az Economist is  Nem, nem Tolkien mester egy eddig kallódó, de most hirtelen fellelt művéről lesz itt szó, hanem a Flores szigetén nemrég fellelt
Nem, nem Tolkien mester egy eddig kallódó, de most hirtelen fellelt művéről lesz itt szó, hanem a Flores szigetén nemrég fellelt 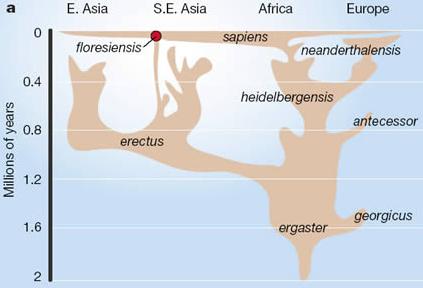 Ugyanakkor már a felfedezés óta hallhatóak szkeptikus hangok, akik szerint egyáltalán nincs új fajról szó, egyszerűen egy pigmeus törzs egyik egyedére, vagy a véletlen folytán egy
Ugyanakkor már a felfedezés óta hallhatóak szkeptikus hangok, akik szerint egyáltalán nincs új fajról szó, egyszerűen egy pigmeus törzs egyik egyedére, vagy a véletlen folytán egy 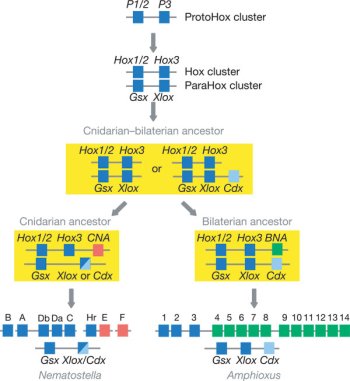 Ugyanis a csalánozó genomok segítségével rekonstruálhatjuk a Bilateria-Cnidaria csoportok szétválása előtti helyzetet (egyébként lásd
Ugyanis a csalánozó genomok segítségével rekonstruálhatjuk a Bilateria-Cnidaria csoportok szétválása előtti helyzetet (egyébként lásd  Nem mehetek el amellett szónélkül, ha a világ második legokosabb élőlényét gyalázzák ;-), már csak azért sem, mert a
Nem mehetek el amellett szónélkül, ha a világ második legokosabb élőlényét gyalázzák ;-), már csak azért sem, mert a