

Az evolúció nagy lépéseinek egyike (legalábbis az evolúciós fa egyes ágain) a többsejtűség kialakulása volt. Előnyei nem igen vitathatóak – hiszen a több sejt nagyobb munkamegosztást tesz lehetővé, amely révén összetettebb feladatok oldhatóak meg -, ugyanakkor még ma is keveset tudunk arról, hogy pontosan milyen mechanizmusok tették lehetővé megjelenését.
A többsejtű szervezetek megjelenésének egyik legfontosabb velejárója, hogy már a fejlődés kezdeti szakaszában elkülönülnek a testi, avagy szomatikus sejtek a szaporodásban később résztvevő ivarsejtektől. Kicsit sarkítva akár úgy is fogalmazhatunk, hogy a szomatikus sejtek önzetlen beáldozzák szaporodási képességüket, a (sejt)közösség, azaz az élőlény szaporodási sikerének maximalizálása érdekében.
Most páran nekiláttak megvizsgálni, hogy miként történik ez a szomatikus-ivari szerepfelosztás egészen egyszerű többsejtes szervezetek esetében. A választás a klasszikus tankönyvi példára a zöld alga sejtekből álló labdára emlékeztető Volvox carteri-re esett, amely több egysejtű zöldalgával együtt a Volvocales csoporthoz tartozik.
A V. carteri mintegy 2000 sejtjéből mindössze 16 ivarsejt, ezek a fejlődés során a hatodik sejtosztódás során jönnek létre, amely asszimetrikus utódsejteket eredményez. A kisebb szomatikus őssejtek aztán még vagy 10-11x újból osztódnak, és végül csillókkal ellátott sejtekké differenciálódnak, amelyek elősegíti, hogy a Volvox mindig a számára optimális megvilágítású helyen lebegjen a vízben.
Mint korábban kiderült, a szomatikus sejtdiffrenciálódás szükséges és elégséges feltétele a regA nevű gén átíródása. Ez a gén aztán meggátolja a kloroplaszt (a napfényt kémiai energiává alakító sejtszervecske) kialakításához szükséges más gének transzkripcióját, s kloroplasztok hiányában a sejtek nem tudnak növekedni, növekedés nélkül pedig nem tudnak osztódni sem – szöges ellentétben az ivarsejtekkel.
Honnan ered hát ez a szabályozás? Ennek megválaszolására egy egysejtű Volvocalesben, a Clamydomonas reinhartdii-ban is megkeresték a regA megfelelőjét. (Ez nem volt teljesen triviális, mert bár a gén kulcsmotívumai jelen voltak, a két szervezetben a gén elég különbözővé alakult az évmilliók során.) S érdekes módon ez a regA homológ is csak akkor fejeződött ki, amikor az egysejtű alga sötétben volt, vagyis napfény és a belőle származó energia hiányában kénytelen volt saját osztódó mechanizmusát kikapcsolni!
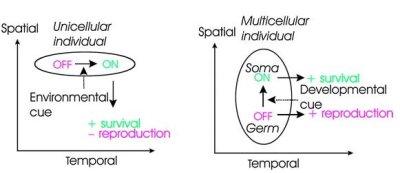
Összefoglalva, úgy tűnik, hogy nem történt más, mint egy már meglevő mechanizmus, amely környezeti hatásokra kikapcsolta az osztódást (azaz időben szétválasztotta a sejt két állapotát), új funkciót nyert (szépen mondva kooptálódott) s a szomatikus- és ivari sejtvonalak (térbeli) szétválasztásában kezdett szerepet játszani.
Hogy a Volvox által használt mechanizmus mennyire volt általános a többsejtűség megjelenésében, természetesen még nemigen tudható, de mindenesetre egy ügyes trükköt láthatunk. Ráadásul újból csak megerősíti azt amit már sokszor, sok helyen elmondtak: új tulajdonságok megjelenésében kulcsszerepe volt a már létező, de addig másra használt mechanizmusok "újradrótozásának" (szabályozásuk megváltoztatásának).
Nedelcu, AM, Michod, RE (2006) The Evolutionary Origin of an Altruistic Gene. Mol Bio and Evo 23(8): 1460-1464.