
 Állati szarvakról beszélve (már csak nyelvi okok miatt is) elsősorban mindenkinek a szarvas szokott beugrani, pedig az igazi nagymesterei a szarvnövesztésnek a bogarak között találhatók. A Scarabaeoidea csoportba tartozó hatlábúak közül több ezer „felszarvazott”, s ha egy-egy múzeumban vesszük az időt és elidőzünk az említetteket bemutató üvegek felett, akár úgy is tűnhet, hogy a szarvak méretének, alakjának, helyzetének csak a képzelet szab határt.
Állati szarvakról beszélve (már csak nyelvi okok miatt is) elsősorban mindenkinek a szarvas szokott beugrani, pedig az igazi nagymesterei a szarvnövesztésnek a bogarak között találhatók. A Scarabaeoidea csoportba tartozó hatlábúak közül több ezer „felszarvazott”, s ha egy-egy múzeumban vesszük az időt és elidőzünk az említetteket bemutató üvegek felett, akár úgy is tűnhet, hogy a szarvak méretének, alakjának, helyzetének csak a képzelet szab határt.
A valóság ennél persze bonyolultabb és, bár a párokért folytatott küzdelem során előszeretettel használt testnyúlványok sokszínűsége kétségtelenül lenyűgöző, jól körülvonalazható, hogy milyen fejlődésbiológiai okok állnak a sokszínűség mögött és azt is sejteni lehet (bár még csak nagyvonalakban), hogy milyen genetikai változások vezettek a szarvak megjelenéséhez és fejlődéséhez.
 Már Darwint és kortársait is megragadta és foglalkoztatta, hogy miként jöhettek létre a bonyolult kitinképződmények, s az sem kerülte el a figyelmüket, hogy a már említett lemezescsápú bogarak (Scarabaeoidea) csoportjának, mintha „különleges hajlama” lenne ezen képződmények létrehozására.
Már Darwint és kortársait is megragadta és foglalkoztatta, hogy miként jöhettek létre a bonyolult kitinképződmények, s az sem kerülte el a figyelmüket, hogy a már említett lemezescsápú bogarak (Scarabaeoidea) csoportjának, mintha „különleges hajlama” lenne ezen képződmények létrehozására.
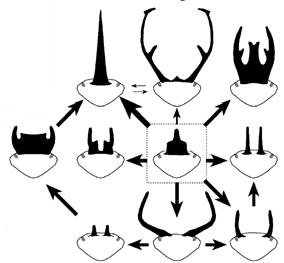
Bár fosszilis ősi skarabeus hiányában nehéz biztosat mondani, ma a kutatók jelentős része úgy gondolja, hogy ez a „különleges hajlam” arra vezethető vissza, hogy a csoport közös őse feltehetően maga is rendelkezett szarvval, s utódai ezt az ősi szarvfejlődési programot módosítgatták, illetve esetenként (pl. a szarv nélküli lemezescsápúak) akár teljesen kikapcsolták. (A baloldali ábra különböző szarvtípusok gazdáinak rokonsági kapcsolatai alapján ábrázolja, hogy feltehetően milyen átmenetek jöttek létre az évmilliók során.) Az átalakulások egyik mozgatóereje sokak szerint a szexuális szelekció lehetett, s mint hamarosan látni fogjuk a nemi vonatkozások visszatérő szereplői a bogárszarvak történetének.
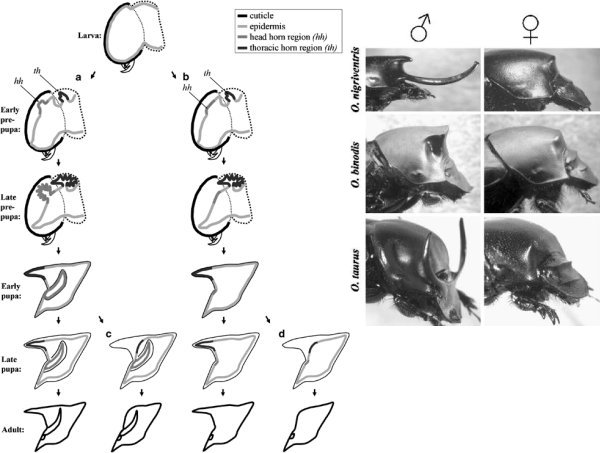
Alapvetően két szarvtípust különböztethetünk meg: konkrétan a fejen előforduló (páros) szarvakat, illetve a torból eredő (gyakran páratlan) szarvakat. Mindkét típusú szarv fejlődésének első fontos szakasza a lárvákban zajlik le, méghozzá meglehetősen gyors ütemben. Mindössze két nappal a báb stádium kezdete előtt (ezt nevezik prepupa szakasznak is) a lárva külső védőrétege, a kutikula alatt az hámszövet (epidermisz) egyes sejtcsoportjai gyors osztódásba kezdenek. Mivel a merev kutikula levedléséig igazából az állat helyszűkében van, a sejtosztódás során létrejövő szervek gyakorlatilag a szó szoros értelmében begyűrődnek a kutikula alá, s csak a lárva stádium végén bekövetkező vedléskor lesz alkalmuk kinyúlni (ezt úgy kell elképzelni, mint amikor egy hosszú lufiba hirtelen levegőt fújunk, s az addig gyűrött kis műanyag darabból hirtelen egy hurka alakú henger lesz). Ekkor aztán az epidermisz sejtjei (amelyek egyébként korábban maguk hozták létre váladékukból a lárva kutikuláját) által termelt anyag a levegőn ismét megszilárdul, és létrehozza a báb külső védőrétegét, amely ezen fejlődési szakasz végéig hivatott óvni a fejlődő bogarat.
A szarvak végső alakja szempontjából kritikus második fázis már a bábokban játszódik le. Ekkor már nem a növekedésen van a hangsúly, sőt sok esetben, feltehetően programozott sejthalállal, a torból eredő szarvak el is sorvadhatnak (lásd a következő ábra, bal oldali részének c. és d. pontját). Ez utóbbi bekövetkezhet mindkét nemben (ez egyébként a ritkább), illetve csak a nőstényekben és a kisebb méretű hímekben, ezzel egyfajta szexuális kétalakúságot (dimorfizmust) hozva létre (erre jó példa a lent látható három Onthophagus faj). A dimorfizmus persze létrejöhet amiatt is, hogy már eleve ki sem alakul egyik vagy másik szarv a lárvában (lásd alábbi ábra, bal oldali részén a b.-t), de bárhogy is van, nem elhanyagolható kérdés, hogy mi az oka annak, hogy egyik nemben kialakul a jelleg a másikban pedig nem, vagy méginkább, hogyan lehet, hogy egy fajban adott nemen belül sem mindenki lesz a szarv büszke hordozója.
Mielőtt azonban tovább mennénk a szexuális dimorfizmus eredetét megpiszkálni, álljunk meg egy pillanatra és próbáljunk meg arra válaszolni, hogy milyen genetikai mechanizmusok hozzák létre a lárvákban a szarvkezdeményeket. Érdekes módon ez egy olyan eset, amikor a gyermeki naivitás adhatja a legjobb tippet. Képzeljük el egy orrszarvú skarabeus elnagyolt plüssváltozatát. Ha elvonatkoztatunk attól, hogy a kezünkben levő valami egy állatot ábrázol, akkor azt mondhatjuk, hogy egy hengeres valamiből lóg ki 11 különböző méretű másik hengeres valami: 2 pár szárny, 3 pár láb ill. maga a szarv. S mint kiderült, a rovarok esetén valóban szinte ilyen „egyszerű” ez: az egyes függelékek ugyanis nagyon hasonló módon alakulnak ki.
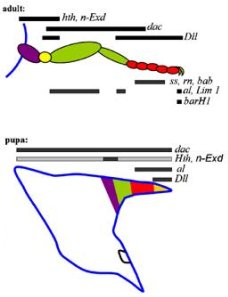 A fejlődésük egy-egy ún. imágó-lemeznek (imaginal disc) nevezett sejtcsoportból indul ki, amely szinte autonóm képződményként működik: a test legváltozatosabb részeibe ültetve át képes végigfuttatni az eredeti programját s létrehozni pl. egy lábat. A lemezek kialakiításáért a wingless (wg) és decapentaplegic (dpp) gének felelősek, az ő termékeik aktiválják majd a lemez területén a függelék tagolásáért felelős többi gént. Ha fentről nézünk az imágó-lemezre, akkor ez utóbbiak expressziója koncentrikus körként jelentkezik. A prepupa szakasz gyors sejtosztódásainak idején aztán a lemez közepéből alakul ki a függelék távoli (disztális) vége, míg a lemez széleiből értelemszerűen a függelék testfalhoz/fejhez ízesülő (proximális) része. (Visszatérve a lufis hasonlatra, képzeljük el, hogy a nem felfújt lufit kifeszítjük és színes köröket rajzolunk rá – ezek aztán a felfújt lufiban hosszanti sávokként jelennek meg.)
A fejlődésük egy-egy ún. imágó-lemeznek (imaginal disc) nevezett sejtcsoportból indul ki, amely szinte autonóm képződményként működik: a test legváltozatosabb részeibe ültetve át képes végigfuttatni az eredeti programját s létrehozni pl. egy lábat. A lemezek kialakiításáért a wingless (wg) és decapentaplegic (dpp) gének felelősek, az ő termékeik aktiválják majd a lemez területén a függelék tagolásáért felelős többi gént. Ha fentről nézünk az imágó-lemezre, akkor ez utóbbiak expressziója koncentrikus körként jelentkezik. A prepupa szakasz gyors sejtosztódásainak idején aztán a lemez közepéből alakul ki a függelék távoli (disztális) vége, míg a lemez széleiből értelemszerűen a függelék testfalhoz/fejhez ízesülő (proximális) része. (Visszatérve a lufis hasonlatra, képzeljük el, hogy a nem felfújt lufit kifeszítjük és színes köröket rajzolunk rá – ezek aztán a felfújt lufiban hosszanti sávokként jelennek meg.)
A láb és szarv imágó-lemezek hasonlósága azt is jelenti, hogy hasonló gének fejeződnek ki bennük, hasonló sorrendben. A jobb oldalon levő ábra azt mutatja be, hogy a szarvakat ugyanazok a gének tagolják, mint a rovarok lábát általában – ha nem is azonos, de nagyon hasonló expressziós mintával. Azaz elég kézenfekvőnek tűnik, azt mondani, hogy a skarabeusok evolúciója során feltehetőleg a korábban a végtagok alakítgatására szolgálódott jelátviteli mechanizmus kooptálódott, vagyis új szerepet kapott a szarvfejlődés során.
A kis embriológiai kitérő után lássuk akkor, hogy mi van a szexuális dimorfizmus hátterében. Úgy tűnik, ebből a szempontból a bogarak sem különböznek tőlünk, hiszen itt is a hormonok okozzák a különbségeket. A kulcs szereplők az inzulin illetve a juvenális hormon (JH).
Mint sok más fajban a korai fejlődés körülményeinek nagy kihatása lesz a felnőtt bogarakra: általában az optimális körülmények között felnnövő, sok táplálékot kapó lárvákból nagy szarvú, domináns hím lesz, míg pechesebb társaikból, szubdomináns szarvatlan hím alakul ki. A táplálkozás indirekt módon hozzájárul az inzulin szint szabályozásához: a megfelelő táplálkozás hatására megnő a hormon koncentrációja, ami a szarvak dinamikus növekedését okozza. A rosszul táplált hímekben (illetve a nőstényekben általában) ezzel szemben az alacsony inzulin szint következményeként elmarad a sejtosztódás. Az inzulinhoz hasonló hatású a JH is, ráadásul ez utóbbi arról híres, hogy a rovarokban megfigyelt más evolúciós változásért is felelőssé volt tehető (itt sincs másképpen s erre hamarosan vissza is térünk).
Az, hogy a nagy hímek számára milyen evolúciós haszonnal bír a nagy szarv az könnyen belátható: akárcsak a szarvasok esetén segít a nőstényekért vívott ádáz csatákban. De miért mondatta le az evolúció a kis hímeket teljesen a szarv növesztésről. A válasz az is lehetne, hogy hasonlóan a hangyák dolgozóihoz, ők is szorgos, de nem szaporodó tagjaivá válnak populációjuknak. De a helyzet koránt sem ez, hiszen ezek a bogarak nem élnek altruisztikus társadalmakban és ennek megfelelően a kis hímek sem mondanak le teljesen a gyarló testi örömökről.
 Ugyanis megfelelő körülmények között a szarvnélküliség akár áldás is lehet, és az ezen bogarak által lakott földalatti járatrendszerek ilyen körülményeknek tűnik. A szűkebb járatokban sokkal könnyebben lehet mozogni nagy szarvak nélkül, s habár környezetük domináns híme ellen szemtől-szemben esélyük sem lenne, a járatokban ide-oda somfordálva azért a kis hímek is el-el csíphetnek egy-egy unatkozó nőstényt. Ez persze azt is jelenti, hogy különböző körülmények között élő populációk esetében esetleg más és más lehet a szarvas/szarvtalan hímek optimális aránya. Ha relatíve kevés hím van a környéken akár mindegyiknek lehet szarva, ha pedig sok akkor pont, hogy jobb kétszer meggondolni mielőtt valaki ilyesmibe fektet.
Ugyanis megfelelő körülmények között a szarvnélküliség akár áldás is lehet, és az ezen bogarak által lakott földalatti járatrendszerek ilyen körülményeknek tűnik. A szűkebb járatokban sokkal könnyebben lehet mozogni nagy szarvak nélkül, s habár környezetük domináns híme ellen szemtől-szemben esélyük sem lenne, a járatokban ide-oda somfordálva azért a kis hímek is el-el csíphetnek egy-egy unatkozó nőstényt. Ez persze azt is jelenti, hogy különböző körülmények között élő populációk esetében esetleg más és más lehet a szarvas/szarvtalan hímek optimális aránya. Ha relatíve kevés hím van a környéken akár mindegyiknek lehet szarva, ha pedig sok akkor pont, hogy jobb kétszer meggondolni mielőtt valaki ilyesmibe fektet.
Az ökológiailag elég tüzetesen vizsgált Onthophagus taurus esetében éppen ezt találták: az eredetileg a mediterrán térségben élő rovart az hetvenes évek környékén telepítették be Ausztráliába illetve az Egyesült Államok keleti részébe. A két új helyszínen a populációk azonban gyökeresen ellenkező módon kezdtek átalakulni. Míg az amerikai O. taurus-oknál a kisebb hímek is elkezdtek szarvat növeszteni, addig a kenguruk földjén élő rokonaiknál már csak a legnagyobb hímek rendelkeznek ilyen függelékekkel.
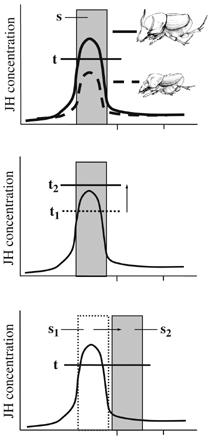
Az ilyen típusú evolúciós változások viszonylag egyszerűen megmagyarázhatók a JH koncentráció segítségével. A hormon koncentrációja egy kritikus értéket kell elérjen (t) egy olyan időszakban (s), amikor a fejlődő állat sejtjei képesek ezt érzékelni, s ha ez bekövetkezik, az állat szarvat növeszt. Ha azonban a kritikus érték megnő, vagy lecsökken (azaz a hormon hatására aktivált gének érzékenysége megváltozik), illetve ha a kritikus periódus „ablaka” elcsúszik, az azzal jár, hogy a környéken rohangáló felszaravazott hímek aránya más lesz, mint a kiindulási populációban lett volna, hasonló körülmények között.
Na, akkor végül összefoglalóban annyit, hogy ez a (terveimnél lényegesen hosszabra nyúlt) poszt arra mutat jó példát, hogy a szervezetben már jelen levő mechanizmusok szabályozásának megpiszkálásával az evolúció nemcsak új jegyeket „varázsolt” a bogarak egy csoportjára, de azokat a szükségnek megfelelően a természetes szelekció révén variálni is képes.
Moczek, AP (2006) Integrating micro- and macroevolution of development through the study of horned beetles. Heredity 97: 168-178.
Emlen, DJ, Szafran, Q, Corley, LS, Dworkin, I (2006) Insulin signaling and limb-patterning: candidate pathways for the origin and evolutionary diversification of beetle ‘horns’. Heredity 97: 179-191.
Kérdésen volna: Az a laikus benyomásom, hogy a mandibula nagy mérete (a szarvasbogáré) és a tor- és fejfüggelékek nagy mérete között van összefüggés: a lemezescsápóak hajlamosak arra, hogy azon a vidéken nagy dolgokat növesszenek. Valóban van ilyen összefüggés?
Teljesen oszinten passz. Ugyanakkor mivel szerintem a szarvak merete sokkal szelesebb intervallumban valtozik mint a csapragoke, ha van is ilyen korrelacio az biztosan nem egyszeru.
Hello Dolphin!
Különösképpen megörvendeztettél ezzel a postoddal (bogarak is, meg fejlődésbiológia is 🙂 …)! Köszi, köszi!
arra reagálva, amit miki is felvetett: valószínűleg pont ez a lényeg, mármint, hogy összefüggés van a mandibula (végtagszármazék), és a fej-, ill. torfüggelékek (nem-végtagszármazék) mérete között. vagyis gyanús, hogy a korai embrionális mintázatképző folyamatokban (hh, wg…) és/vagy a későbbi proliferációs szakaszban (inzulin…) lehet valami a lemezescsápúakban, ami nincs más bogárcsaládoknál (ez azért nem teljesen igaz). tehát lehet hogy nem is annyira a „szarv” a közös fenotipikus jelleg a csoporton belül, hanem az azt kialakító molekuláris mintázatképző folyamat egyik eleme. ezt megtalálva talán a közös ős is jobban körvonalazható lesz.
mint ahogyan a cikk is megjegyzi, ezeknél a bogaraknál nagyon jól megkülönböztethetők a „domináns” és „szubdomináns” fenotípusú egyedek (régebben a rendszerezők szerették ezeket külön változatnak tekinteni, meg elnevezgetni). ez arra utal, hogy van egy „mindent vagy semmit” lépés a fejlődésükben: valószínüleg épp a JH-nak kell elérnie egy kritikus szintet ahhoz, hogy a sejtek proliferációja beinduljon, és így a függelékek kialakuljanak.
+1 megjegyzés: csáprágójuk a pókoknak van 🙂
Dolphin! Hogy lehet neked e-mailt írni? Szeretnék neked új cikket ajánlani. Néha belebotlok olyan publikációba, hírbe, ami pont ide illene, legalább elküldeném neked, hátha megtetszik…
Emil cim: dolphinm kukac gmail pont com.
Nem igerek biztosra semmit, mert az idom az nagyon szuk keresztmetszet, de azert majd igyexem :-).