 A külalak (pl. szőrzetszín) változása mindig hálás témája az evolúciót taglaló irományoknak, hiszen az még a leglaikusabb olvasók számára is triviális, hogy mind a ragadozók, mind a prédáik számára nagyon fontos, hogy minnél jobban kerüljék a feltűnést és beleolvadjanak környezetükbe.
A külalak (pl. szőrzetszín) változása mindig hálás témája az evolúciót taglaló irományoknak, hiszen az még a leglaikusabb olvasók számára is triviális, hogy mind a ragadozók, mind a prédáik számára nagyon fontos, hogy minnél jobban kerüljék a feltűnést és beleolvadjanak környezetükbe.
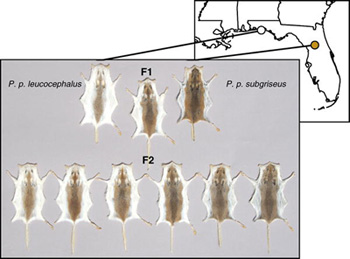
Nincs ez másként egy floridai egérfaj, a Peromyscus polionotus esetében sem, ahol a homokos part közelében élő populációk, lényegesen világosabb, homokszínű bundával rendelkeznek, mint a félsziget belsejében elő társaik. A jelleg adaptív voltához nem sok kétség férhet, s mivel sok más emlősfajnál (pl. mamutok) a hasonló adaptív változásokat a melanocortin-1-receptor (mc1r) változásához lehetett kapcsolni, ez esetben is először ezt a gént vizsgálták meg a kutatók.
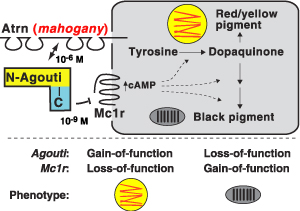
Normális körülmények között az Mc1R feladata, hogy a melanocortin stimuláló hormon (msh) hatására instruálja a sejteket sötét pigment, azaz eumelanin termelésére. Ha azonban egy olyan mutáció jelenik meg a génben, ami csökkenti az aktivitását, az értelemszerűen világosabb mintázat formájában fog jelentkezni az állat hétköznapjaiban.
 Pont egy ilyen mutációra leltek a P. polionotus parti populációiban (P. p. leucocephalus) is, amikor szúrópróbaszerűen megvizsgálták az mc1r gént, de ugyanakkor az is gyanítható volt, hogy a mutáció nem egyedül felelős a világosabb szőrzetért.
Pont egy ilyen mutációra leltek a P. polionotus parti populációiban (P. p. leucocephalus) is, amikor szúrópróbaszerűen megvizsgálták az mc1r gént, de ugyanakkor az is gyanítható volt, hogy a mutáció nem egyedül felelős a világosabb szőrzetért.
Ezért a következő lépésben a Hopi Hoekstra vezette csoport szisztematikusabb vizsgálódásba kezdett. A tüskés pikós posztban már körülírt módszert alkalmazva, keresztezték a part menti világos és a parttól távól élő sötét egérközösségek képviselőit, és azt nézték, hogy a világosabb bunda szín melyik genetikai markerekkel öröklődik együtt (ez ugyanis arra utal, hogy a marker közelében levő génnek szerepe lehet az új jelleg kialakulásában). A lista viszonylag rövid lett, mindössze három gén szerepelt rajta (és egyik sem „új”, olyan szempontból, hogy a laboratóriumi egerekkel végzett kísérletek során, már kapcsolatba hozták a szőrzet pigmentációjával): az első – nem túl meglepő módon – az mc1r, a második egy agouti nevű gén, a harmadik pedig a c-kit receptor (Kit). A vizsgálatok szerint a három közül az első kettőnek van igazán fontos szerepe, így bár a Kit működése sok szempontból érdekes, a továbbiakban hanyagolták.
Az agouti a pigmentációs folyamatokban az Mc1r antagonistájaként működik, azaz hozzá kapcsolódva gátolja a működését – így ahol jelen van nem, vagy csak alig keletkezik sötét pigment. A floridai parti egerek bőrében az agouti kifejeződési szintje mindenhol erőteljesen megemelkedett, s így kombinálódva a kevésbé aktív Mc1r receptorral együttesen „fehérítik ki” a mindkét mutációt hordozó egereket. (Bár, tegyük hozzá, csupán az egyik mutáció birtoklása is világosabb szőrzetet eredményez – az igazán hasznos, „homokszínhez”, azonban mindkettő kell.)
Vagyis a Mexikói öböl bícsein héderező rágcsálók egyszerre jó példái annak, hogy az evolúció és a természetes szelekció egyaránt működhet gének funkcióinak elvesztésével, vagy korlátozásával (lásd mc1r), és új funkciók megjelenésével, vagy már meglevők kiterjesztésével (lásd agouti). Mikor épp miből lehet előnyt kovácsolni.
Steiner CC, Weber JN, Hoekstra HE (2007) Adaptive Variation in Beach Mice Produced by Two Interacting Pigmentation Genes. PLoS Biol 5(9): e219 doi:10.1371/journal.pbio.0050219

 Indiánokról írni általában véve hálás feladat, hiszen a téma természeténél fogva, mindig akad egy kis egzotikum, vadregényes tájak, számunkra furcsa kinézetű és viselkedésű emberek. És mint nagy írónk ;-)) May Károly esete is bizonyítja, még csak el sem kell utazni az amerikai kontinensre, hogy a témáról bestsellereket írjunk.
Indiánokról írni általában véve hálás feladat, hiszen a téma természeténél fogva, mindig akad egy kis egzotikum, vadregényes tájak, számunkra furcsa kinézetű és viselkedésű emberek. És mint nagy írónk ;-)) May Károly esete is bizonyítja, még csak el sem kell utazni az amerikai kontinensre, hogy a témáról bestsellereket írjunk. Úgy tűnik, hogy hirtelen nagyon trendi lett neandervölgyi géneket vizsgálni, hiszen
Úgy tűnik, hogy hirtelen nagyon trendi lett neandervölgyi géneket vizsgálni, hiszen 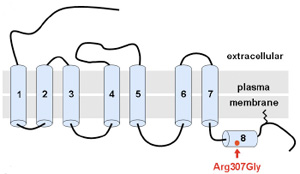 A neandervölgyi mc1r különlegessége, hogy bizonyos esetekben a jelek szerint egy olyan jellegzetes mutációt hordoz(ott), aminek a következtében, működésbe lépésekor a cAMP koncentráció nem éri el azt a szintet, amit a sötét hajú emberekben figyelhetünk meg. Érdekes módon, ez nem azért van (volt), mert a receptor kevésbé hatékony, hanem mert (pontosan nem tisztázott okok miatt) eleve kevesebb mutáns receptorfehérje jut el a sejtek belsejéből a felszínükre. A végeredmény szempontjából azonban ez mindegy és a létrejövő alacsonyabb cAMP szint megegyezik azzal, amit vöröshajú emberekből izolált MC1R hoz létre – azaz jó eséllyel feltételezhető, hogy az ősi neandervölgyiek kb. 1%-a vörös fej- és arcszőrzettel büszkélkedhetett.
A neandervölgyi mc1r különlegessége, hogy bizonyos esetekben a jelek szerint egy olyan jellegzetes mutációt hordoz(ott), aminek a következtében, működésbe lépésekor a cAMP koncentráció nem éri el azt a szintet, amit a sötét hajú emberekben figyelhetünk meg. Érdekes módon, ez nem azért van (volt), mert a receptor kevésbé hatékony, hanem mert (pontosan nem tisztázott okok miatt) eleve kevesebb mutáns receptorfehérje jut el a sejtek belsejéből a felszínükre. A végeredmény szempontjából azonban ez mindegy és a létrejövő alacsonyabb cAMP szint megegyezik azzal, amit vöröshajú emberekből izolált MC1R hoz létre – azaz jó eséllyel feltételezhető, hogy az ősi neandervölgyiek kb. 1%-a vörös fej- és arcszőrzettel büszkélkedhetett. Ha kellően hiszékenyek vagyunk, akkor
Ha kellően hiszékenyek vagyunk, akkor  Az 1950-es évek közepén, miközben a biológusok világszerte csak a
Az 1950-es évek közepén, miközben a biológusok világszerte csak a  Ha a rovarvilág molekuláris svájcibicskáját keresnénk, az Ubx gén jó eséllyel pályázhatna erre a címre. Szerepe van abban, hogy a rovarok potrohán
Ha a rovarvilág molekuláris svájcibicskáját keresnénk, az Ubx gén jó eséllyel pályázhatna erre a címre. Szerepe van abban, hogy a rovarok potrohán  Mint az alig pár hete
Mint az alig pár hete 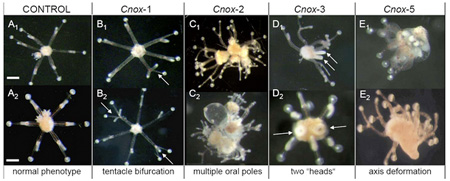 A Hox-gének,
A Hox-gének, 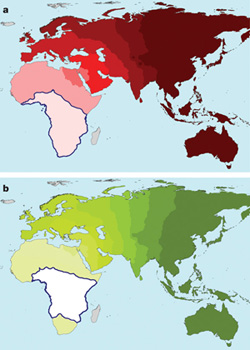 Akik járatosak a témában, biztos felfigyeltek rá, hogy mikor a múlt héten az emberi sokféleség eredetét (pontosabban amit erről ma tudunk/gondolunk)
Akik járatosak a témában, biztos felfigyeltek rá, hogy mikor a múlt héten az emberi sokféleség eredetét (pontosabban amit erről ma tudunk/gondolunk)