
Ahogy azt még a blog hajnalán írtam egyszer, a világ legősibb sejtvonala nem valamilyen sztárlabor inkubátorainak mélyén tenyészik Petri-csészékben, hanem hús-vér kutyák segítségével maradt fent, több ezer éven át.

Ahogy azt még a blog hajnalán írtam egyszer, a világ legősibb sejtvonala nem valamilyen sztárlabor inkubátorainak mélyén tenyészik Petri-csészékben, hanem hús-vér kutyák segítségével maradt fent, több ezer éven át.

Nyolc évvel ezelőtt (jönne, hogy írjam „pár éve”, pedig dehogy pár éve az…) már írtam egyszer egy különleges mutációról, ami egyes kutyafajták esetében (pl. tacskók és bassett houndok) rövid végtagok kialakulását eredményezi. Azért volt különleges, mert nem egyszerűen egy már meglevő gén aminosavsorrendje, vagy szabályozása változott meg, hanem egy génnek egy újabb kópiája keletkezett. Mégpedig nem genomduplikációval, hanem egy reverz transzkriptáz enzim segítségével az eredeti, genomi DNS-ből átíródott (és a splicing során megérett) FGF4 mRNS íródott vissza (immár intronok nélküli) DNS darabbá és integrálódott a genomba a 18. kromoszómán.
Azt gondolnánk, hogy egy ilyen esemény már önmagában is elég különleges, ám pont a napokban arra derült fény, hogy szinte hajszálpontosan ugyanez az esemény még egyszer lejátszódott a kutyák tenyésztése során.

A farkasok domesztikációja az őstől nem pusztán fizikai jellegükben különböző eb populációkat hozott létre, hiszen a különböző kutyafajták közös jellemzője, hogy viselkedésükben is jellegzetes módon eltérnek az ősi fajtól.
 A kutyák farkas-eredete tulajdonképpen nem képezi semmilyen vita tárgyát, leginkább az esemény pontos „hol” és „mikor”-ja kérdéses.
A kutyák farkas-eredete tulajdonképpen nem képezi semmilyen vita tárgyát, leginkább az esemény pontos „hol” és „mikor”-ja kérdéses.
Korábbi eredmények kb 15 000 évvel ezelőttre tehető a kutya és farkas populációk szétválásának ideje, valahol Eurázsiában. A vizsgálatot természetesen nehezíti, hogy a két közösség nem csak hogy elméletileg képes szaporodni egymással, hanem a világ számos pontján az emberek aktív segítségével teszik ezt, hiszen ez mindig is a kutya-vérvonal „frissítésének” egyik titka volt.
Márpedig az ilyen eseti együttlétek bonyolítják a két fajta szétválásának dekódolását a genomjaik alapján.
 Kevés faj rendelkezik olyan formagazdagságal, mint a kutya (pontosabban a szürke farkas, hiszen ő a faj vadon élő ősi képviselője), az emlősök közül pedig jó eséllyel senki.
Kevés faj rendelkezik olyan formagazdagságal, mint a kutya (pontosabban a szürke farkas, hiszen ő a faj vadon élő ősi képviselője), az emlősök közül pedig jó eséllyel senki.
A hatalmas formagazdagság elsősorban az emberi tenyésztők szorgalmát és néha jobb, néha furább ízlését dicséri, ami azzal a bónusszal is jár, hogy számos kutya fajta esetében nagyon részletes családfák állnak a rendelkezésünkre, ami mindig is megkönnyítette az egyes fajták különleges tulajdonságaiért felelős gének feltérképezését. Persze napjaink modern és gyors szekvenálási eljárásai még rátettek minderre egy lapáttal, így az utóbbi években szinte csak özönlöttek a külünböző kutyagenetikával fogallkozó cikkek a legrangosabb lapokban is. A kutatásban Elaine Ostrander csoportja járt az élen (lásd még eredményeikről itt és itt és itt) és a PLoS Geneticsben nemrég megjelent cikkük is ebbe a sorba illik bele.
 Mint azt két hete már beígértem, most az Ostrander laboratórium egyik legutóbbi eredményéről lesz szó, amiben a különböző kutya szőrtípusok genetikai eredetének erednek nyomába.
Mint azt két hete már beígértem, most az Ostrander laboratórium egyik legutóbbi eredményéről lesz szó, amiben a különböző kutya szőrtípusok genetikai eredetének erednek nyomába.
Kettős megközelítést használtak: egyrészt azt figyelték, hogy léteznek-e olyan genetikai markerek, amelyek hosszú szőrű/ drótszőrű/ pofaszakállás fajták egyedeiben szinte kivétel nélkül jelen vannak, míg más fajtákban nagyon ritkák. Másrészt kiemelten foglalkoztak egy olyan fajtával, ahol mindezen szőrzet jegyek egyaránt megfigyelhetők: a tacskókról van szó.
Az eredmények egyik érdekessége, hogy a legváltozatosabb fajok esetében ugyanazokat a mutációkat tudták az említett szőrzetjegyekért felelőssé tenni, a másik pedig az, hogy több, megvizsgált farkaspopulációból ezek a mutáns allélvariánsok egyaránt hiányoztak, vagyis a mutációk az ebszelídítés során alakultak ki, és nem pusztán már eredendően, egy ősi populációban meglevő genetikai sokszínűségről van szó, amelyből az emberi ízlés választott ki aztán valami neki tetszőt.
Ha valaki szeret elgondolkozni a kutyavilág sokféleségén, akkor érdemes a fejébe vésnie Elaine Ostrander nevét, mivel a hölgy (és laboratóriuma) jelenleg igencsak bő forrása a legkülönbözőbb jellegek genetikai térképezésének. Az ő nevéhez fűződik, a kis eb-termet és az IGF1 gén adott allélja közti összefüggés megállapítása, a végtaghossz-különbözőségek fejlődési okának beazonosítása (jelen poszt témája), illetve a különböző szőrzettípusok genetikájának tisztázása (erről, remélem, hamarosan).
A rövid végtaggal rendelkező kutyák (pl. tacskók, bassett houndok) esetében, az már régóta világos volt, hogy egy örökletes fejlődési rendellenességről van szó: valamilyen oknál fogva azok a sejtek, amelyek a csont növekedését kellene szolgálják, idő előtt befejezik az osztódást és maguk is „elcsontosodnak”. (A deformált és rövid végtagok jelenlétének klinikai neve chondrodysplasia.) A miértre azonban eddig nem tudtuk a választ.
Ostranderék egy hatalmas adathalmazt gyűjtöttek össze, több száz kutya genomját (és a benne levő genetikai polimorfizmusokat) képesek megvizsgálni rövid idő alatt, aminek oroszlánrésze van abban, hogy viszonylag gyorsan tudnak érdekes tulajdonságoknak a genetikai okaira rákeresni.
Számos rövid ill. hosszú lábú eb fajta összehasonlításából aztán kitűnt, hogy a jelleg nagyon is kapcsolódik a 18. kromoszóma egy adott részén felfedezhető polimorfizmusokhoz, ami kevésbé szakzsargonban azt jelenti, hogy valahol ezek mellett van az a mutáció, amihez a jelleg köthető.
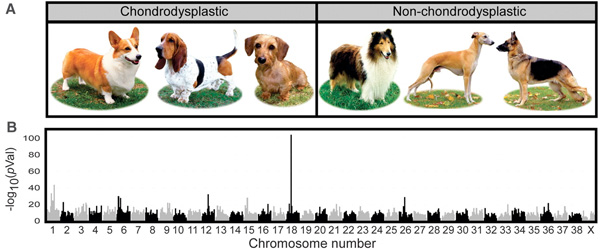
Érdekes módon, a standard kutyagenomban nem nagyon lehet fellelni egyetlen izgalmas gént sem a közelben, aminek egészen prózai oka van: nem egy „tradicionálisan” itt levő génben van a mutáció.
A chondrodypláziás kutyák esetében egy hívatlan vendég jelent itt meg: egy fgf4 retrogén. Mit is jelent ez: az fgf4 normál esetben egy négy exonból és három intronból álló gén (érdekes módon szintén a 18. kromoszómán van, de valahol nagyon máshol, mint a vizsgált régió). Az intronok aztán a transzkripció során/után kivágódnak, és így jön létre a bő 600 bázispár hosszúságú mRNS, ami aztán a fehérje szintézissorán lefordítódik. A vizsgált retrogén azonban egyáltalán nem rendelkezik intronokkal: szekvenciája szinte hajszálra megegyezik az érett mRNS-ével, aminek az a prózai oka van, hogy abból keletkezett. Egy véletlennek köszönhetően, egy ivarsejtben, feltehetően a retrotranszpozon aktivitásának köszönhetően jelenlevő reverz transzkriptáz a transzpozon saját genetikai állománya mellett egy fgf4 mRNS molekulát is „visszafordított” DNS-re, visszacsempészve a genomba. Az így keletkező új „retrogén” történetesen olyan szabályozó szekvenciák közelébe került, amelyek a fejlődés egy adott (csontfejlődés szempontjából kritikus) szakaszában aktiválják a gént a csontnövekedésért felelős sejtekben, amely nem kívánt aktivitásnak aztán egyenes következménye a chondrodyspláziás fenotípus.
A megfigyelés nem teljesen meglepő, mert egerekben és emberekben egyaránt az FGF jelátviteli úthoz lehet gyakran kötni a hasonló jellegű betegségeket. Ami azonban nem triviális, hogy a sok, egyébként igencsak különböző, rövid lábú fajtában ugyanaz a ritka mutáció felelős a jelleg megjelenéséért. Ez ugyanis azt jelenti, hogy még a kutyaszelídítés hajnalán jelent meg ez a mutáció a kutyaállományban és azóta is fennmaradt az emberi ízlésnek és gondoskodásnak köszönhetően.
(A borítókép a Wikimedia Commons oldaláról származik.)
Parker HG, VonHoldt BM, Quignon P, Margulies EH, Shao S, Mosher DS, et al. (2009) An expressed fgf4 retrogene is associated with breed-defining chondrodysplasia in domestic dogs. Science 325: 995-998.
 Mióta magunk alakítjuk (egyre hatékonyabban) a körülöttünk levő világot, fokozatosan átalakult a „rátermettség” definíciója is, mind a saját, mind a hozzánk kapcsolódó élőlények viszonylatában.
Mióta magunk alakítjuk (egyre hatékonyabban) a körülöttünk levő világot, fokozatosan átalakult a „rátermettség” definíciója is, mind a saját, mind a hozzánk kapcsolódó élőlények viszonylatában.
A „legrátermettebb” háziállataink már régen nem feltétlenül azok, amelyek a vadonban a legtöbb utódot hoznák létre, hanem amelyeknek önös okainkból kifolyólag, mi kínáljuk fel a tálcán a lehetőséget, hogy szaporodjanak és sokasodjanak.
És ezzel sokszor „átírjuk” a természet szabályait, olyan génvariációk, allélok elterjedését okozva, amelyek egyébként kihalásra lennének ítélve, legyen szó hófehér, de bőrrákra hajlamos lovakról, vagy izmos, de impotens gyanús tehenekről. Vagy éppen apró, szőr és fognélküli kutyákról, mint a kínai meztelen kutya – hiszen nehéz elképzelni azt a természetesen niche-t, amiért egy ilyen lény sikeresen harcba szállna akármivel, akárkivel.
A kutyagenomnak köszönhetően az utóbbi időben felgyorsult a különleges kutyafajták kínézete mögött rejlő gének feltérképezése (lásd pl. itt, itt és itt), s a héten az előbb említett meztelen eb került sorra.
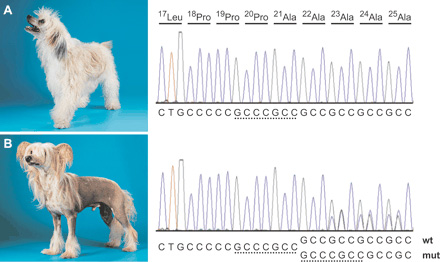
A 17. (kutya)kromoszómára lokalizált jelleg érdekessége, hogy egy monogénikus, szemidomináns jelleg, amely homozigóta formában letális. Konyhanyelvre lefordítva ez annyit jelent, hogy a kopaszság (és fognélküliség) egyetlen génben levő mutációra vezethető vissza, amely csak akkor lesz „látható”, ha a kutya a gén mutáns formája mellett, a hibátlan verziót is hordozza. Ugyanis, ha egy kutyaembrió csak a gén mutáns formájával rendelkezik, akkor a fejlődés korai szakaszában elpusztul.
A jelleget vizsgáló csoport végül egy transzkripciós faktort kódoló génre, a FOXI3-ra szűkítette le a lehetőségek kezdetben széles halmazát. Ez a gén egerekben a szőrtüszőkben és a fejlődő fogakban fejeződik ki, s bár pontos szerepe (vagyis, hogy ő maga, milyen más gének átíródását befolyásolja) még nem tisztázott, mindez értelmet ad a mutáns kutyák kinézetének. Maga a mutáció egy 7 bázispárnyi duplikáció a gén elején (ennek az lehet az oka, lásd a mellékelt ábra, hogy a gén itt GCC bázis -tripletekben gazdag, ami miatt a gén másolásakor, a polimeráz enzim jó eséllyel eltévedhet, s itt egyszer el is tévedt), amelynek következtében egy korai STOP kodon alakul ki, s csak a fehérje első ~5%-a íródhat(na) át.
Hogy a gén szemidomináns jellege miből adódik, az még rejtély. Valószínűleg a fogak és szőrtüszők fejlődéséhez pontosan meghatározott mennyiségre van szükség a FOXI3 fehérjéből, s ha csak az egyik allél van jelen, akkor nem lesz meg ez a kritikus mennyiség.
Drögemüller C, Karlsson EK, Hytönen MK, Perloski M, Dolf G, et al. (2008) A Mutation in Hairless Dogs Implicates FOXI3 in Ectodermal Development. Science 321: 1462.
 Az 1950-es évek közepén, miközben a biológusok világszerte csak a liszenkóizmus abszurditását láthatták a nagy szovjet (tudományos) valóságból, Szibéria egyik szegletében, élettan kísérletnek álcázva, a 20. század egyik legérdekesebb genetikai kísérletét kezdte el, egy félig száműzetésben levő orosz kutató. Dimitrij Beljajev 1948-ban veszette el moszkvai állását, mivel nem volt hajlandó a mendeli genetikát becserélni Liszenkó „micsurini” nézeteire, és rövid kitérő után a Szovjet Tudományos Akadémia novoszibirszki intézetében kötött ki, ahol haláláig dolgozott.
Az 1950-es évek közepén, miközben a biológusok világszerte csak a liszenkóizmus abszurditását láthatták a nagy szovjet (tudományos) valóságból, Szibéria egyik szegletében, élettan kísérletnek álcázva, a 20. század egyik legérdekesebb genetikai kísérletét kezdte el, egy félig száműzetésben levő orosz kutató. Dimitrij Beljajev 1948-ban veszette el moszkvai állását, mivel nem volt hajlandó a mendeli genetikát becserélni Liszenkó „micsurini” nézeteire, és rövid kitérő után a Szovjet Tudományos Akadémia novoszibirszki intézetében kötött ki, ahol haláláig dolgozott.
Beljajevet különösen érdekelte a domesztikáció, az állatszelídítés jelensége és arra figyelt fel, hogy olyan távoli rokon fajok esetében, mint a disznó és a kutya a folyamat hasonló jegyek megjelenésével járt, mint pl. felfele kunkorodó farok, lelógó fülek, megváltozott testszőrszín. Ennek alapján állította fel saját elméletét, mely szerint ezek a jellegek mintegy a szelídítés „melléktermékei” – s ez részben szembement a korabeli kánonnal, mely szerint a domesztikáció során, az ember nem csak egy jegyre, a kezelhetőségre/szelídségre koncentrált, hanem számos másikra is, pl. szociális hajlam, bunda mintázat, koponyaalak, könnyű szaporítás stb.
 A kutyák megunhatatlan alanyai a különböző genetikai vizsgálatoknak, hiszen évezredek szorgos tenyésztésével egészen elképesztő formagazdaságot sikerült létrehoznunk körükben. Így aztán legyen a kérdés méret, vagy alkat, biztos, hogy sikerül valami informatívat „kipréselnünk” négylábú barátainkból.
A kutyák megunhatatlan alanyai a különböző genetikai vizsgálatoknak, hiszen évezredek szorgos tenyésztésével egészen elképesztő formagazdaságot sikerült létrehoznunk körükben. Így aztán legyen a kérdés méret, vagy alkat, biztos, hogy sikerül valami informatívat „kipréselnünk” négylábú barátainkból.
Most épp a whippet (azaz szalonagár) került terítékre. Ezt az agáralkatú kutyát Angliában tenyésztették ki versenyzésre a 17. század magasságában. S bár ma már számos kiállításon a kutya sudár alkatát díjazzák előszeretettel, a verseny-tenyésztők számára a külalak másodlagos, a legfontosabb a sebesség.
{kind=link}