
![]() Az egyik dolog, ami igazán megkülönböztet bennünket emberszabású rokonainktól, az a fajunk által belakott terület sokszínűsége. Természetesen az ember alkalmazkodó képessége elválaszthatatlanul kapcsolódik a szofisztikált eszközhasználathoz, az elvont gondolkozáshoz és komplex kommunikációs képességhez, de a Szahara hőségéhez és a szibériai puszta dermesztő világához való adaptálódás nem merült ki pusztán az öltözködési és étkezési szokások változtatgatásában. A folyamat számos formában rajta hagyta nyomát az évszázadok óta ott élő embertársaink genetikai állományán is – mert a természetes szelekció alól mi sem tudjunk nagyon kivonni magunkat.
Az egyik dolog, ami igazán megkülönböztet bennünket emberszabású rokonainktól, az a fajunk által belakott terület sokszínűsége. Természetesen az ember alkalmazkodó képessége elválaszthatatlanul kapcsolódik a szofisztikált eszközhasználathoz, az elvont gondolkozáshoz és komplex kommunikációs képességhez, de a Szahara hőségéhez és a szibériai puszta dermesztő világához való adaptálódás nem merült ki pusztán az öltözködési és étkezési szokások változtatgatásában. A folyamat számos formában rajta hagyta nyomát az évszázadok óta ott élő embertársaink genetikai állományán is – mert a természetes szelekció alól mi sem tudjunk nagyon kivonni magunkat.
Az adaptáció persze csak az egyik forrása az emberi populációk között észlelhető genetikai különbségeknek, bizonyos esetekben a vakvéletlen legalább ekkora szerepet játszhat egyes gének bizonyos alléljainak elterjedésében/eltűnésében (lásd még a genetikai sodródás jelenségét, amikor a kezdeti sokféleség sztohasztikus hatások nyomán lecsökken). De bármi legyen is az oka a DNS-ben rögzülő változások kialakulásának, egy biztos: azok öröklőd(het)nek és éppen ebből a tulajdonságukból adódóan, ha megfelelően „kérdezzük” őket, mesélni tudnak eredetükről, sőt akár hordozóik ük-ük-…-ük-szüleiről is egy kicsit.
 Két ember genomja átlagosan 3 millió ponton különbözik egymástól, ami elsőre soknak tűnhet, ezért nem árt kontextusba helyezni egy kicsit: 3 milliárd bázispárból álló genomunknak ez mindössze 0.1%-a (azaz egy ezreléke). Hogy ez az állatvilág normái között mennyire kevésnek számít, azt jól szemlélteti, hogy az átlagos genetikai különbözőség még a hozzánk leginkább hasonlító emberszabásúakban is a négyszerese ennek.
Két ember genomja átlagosan 3 millió ponton különbözik egymástól, ami elsőre soknak tűnhet, ezért nem árt kontextusba helyezni egy kicsit: 3 milliárd bázispárból álló genomunknak ez mindössze 0.1%-a (azaz egy ezreléke). Hogy ez az állatvilág normái között mennyire kevésnek számít, azt jól szemlélteti, hogy az átlagos genetikai különbözőség még a hozzánk leginkább hasonlító emberszabásúakban is a négyszerese ennek.
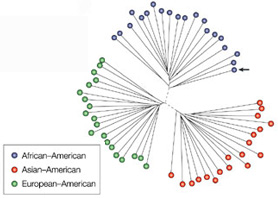
Persze, a 3 millió eltérés azért elég nagy szám ahhoz, hogy leírja egy aborigin bennszülött és egy norvég, vagy egy pigmeus és egy inuit közti kinézetbeli különbséget. Olyannyira, hogy az összes különbség csak mintegy 8%-a (!!) felelős a különböző népcsoportok „tipikus” képviselői közt észlelhető változatosságért, további 8% okolható a populációk különböző csoportjain belül fellelhető különbségekért (pl. átlagban ennyi különbség lesz Sopron és Szeged lakói között), míg a maradék kb. 84% a csoport különböző tagjai (pl. Te, kedves olvasó és a szomszédod) közti különbségre megy el. (Lásd a mellékelt ábrán, a három populáció közti átlagos különbséget jelölő „ágak” középen sokkal rövidebbek, mint a az egyes bokrokon belüli ághosszak.) Ezet az értékeket először Richard Lewontin fedezte fel vércsoportokon végzett kutatásai során, de azóta nagy mintaszámú DNS vizsgálatok is alátámasztották.
S mivel ez az érték sajnos nincs eléggé a köztudatban, nem árt hangsúlyozni. Ugyanis ez azt jelenti, hogy ha valamilyen katasztrófa következtében az emberiség elsöprő többsége elpusztulna, és csak egy eldugott indonéziai törzs maradna meg, vagy egy borsodi község lakossága, a túlélők még mindig a ma élő teljes emberi népesség genetikai diverzitásának 84%-át hordoznák jó eséllyel. Lehetetlenül hangzik? Pedig volt hasonlóra példa (bár kicsit kisebb léptékben) már a történelemben: Dél-Afrika fehérbőrű lakossága gyakorlatilag mind egy szálig kétezer fehérbőrű telepesre vezethető vissza, akik mintegy 350 éve szálltak Capetown környékén partra, vagy ott van a quebeci franciaajkú lakosság, akiknek többsége annak a 770 nőnek a leszármazottja, akik a 17. sz. vége fele, XIV. Lajos szponzorálásával mentek az Újvilágba, hogy az ott élő (nőtlen) telepesek életminőségén lendítsenek egyet. De lehet, hogy a legjobb (és talán legkevésbé ismert) példa Földünk összlakossága, hiszen a korábban emlegetett alacsony genetikai diverzitás sok kutató szerint azzal magyarázható, hogy mintegy szűk 100,000 éve, valamilyen környezeti katasztrófa folytán a teljes H. sapiens populáció alig 10-15,000 egyedre apadt.
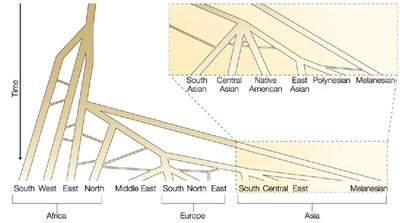 Ennek a pár ezer embernek az utódai lakták be aztán az összes kontinenst (az Antarktisz kivételével), alkalmazkodva a helyi körülményekhez. Nem véletlen, hogy a legtöbb emberi gén, amely úgy tűnik, hogy szelekció alatt van, az immunrendszerhez kapcsolható – hiszen a világ különböző sarkaiban obskurus betegségek, mint a malária, leselkednek az ott élőkre -, vagy éppen a napsugárzás intenzitásához való alkalmazkodást biztosító bőrszínhez. Ez a fokozatos földrajzi terjeszkedés ott is tettenérhető, hogy a szomszédos populációk (falvak, városok) lakói mindig csak kicsit különböznek egymástól; s ez nem csak kontinenseken belül, de köztük is igaz, így aztán nem lehet éles (sőt homályosat se) határt húzni Afrika és Európa, vagy Európa és Ázsia lakói között.
Ennek a pár ezer embernek az utódai lakták be aztán az összes kontinenst (az Antarktisz kivételével), alkalmazkodva a helyi körülményekhez. Nem véletlen, hogy a legtöbb emberi gén, amely úgy tűnik, hogy szelekció alatt van, az immunrendszerhez kapcsolható – hiszen a világ különböző sarkaiban obskurus betegségek, mint a malária, leselkednek az ott élőkre -, vagy éppen a napsugárzás intenzitásához való alkalmazkodást biztosító bőrszínhez. Ez a fokozatos földrajzi terjeszkedés ott is tettenérhető, hogy a szomszédos populációk (falvak, városok) lakói mindig csak kicsit különböznek egymástól; s ez nem csak kontinenseken belül, de köztük is igaz, így aztán nem lehet éles (sőt homályosat se) határt húzni Afrika és Európa, vagy Európa és Ázsia lakói között.
Itt azért az objektivitás okán picit vissza kell térnünk a bizonyos 8%-os különbséghez. Ezt ui. sokan úgy értelmezték, hogy azt jelenti, nagyon nagy az esélye, hogy a Föld tetszőleges két embere, pl. egy dán és egy vietnámi jobban hasonlít egymásra genetikai értelemben, mint a dán és (szintén dán) szomszédja. Ennek tényleg nem elhanyagolható az esélye, de azért kevesebb mint 50% (ami akkor lenne, ha valóban véletlenszerű lenne a dolog), „mindössze” 25-35%, attól függően, milyen csoport képviselőit vizsgáljuk. Ugyanakkor a hasonlóságok elegendőek ahhoz, hogy megkérdőjelezzük van-e értelme a szó klasszikus értelmében „rasszokról” beszélni? Hiszen amit a fogalom megalkotásakor gondoltak a rasszokról, az mind hamisnak bizonyult, s bár kétségtelen, lehetne – és páran meg is próbálnak – összetákolni egy új definíciót, hogy az többé kevésbé földrajzilag lefedje azokat a népcsoportokat, amelyekre fogalom klasszikus értelmezése vonatkozott, vajon nem lenne-e jobb egész egyszerűen arról beszélni, amit valóban vizsgálunk: emberi populációkról? (De, jobb lenne.)
A kis nomenklatúrai kitérő után, térjünk most vissza a sokkal érdekesebb kérdéshez: mi volt a Föld emberi benépesítésének a kronológiája? Hogy ezt vizsgálni tudjuk, a genomunk sajátos részéhez, a nem rekombinálódó DNS-hez kell forduljunk.
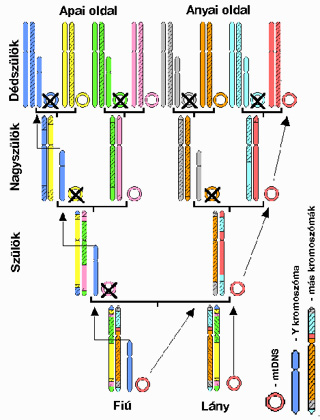 Arra talán mindenki emlékszik gimis tanulmányaiból, hogy a legtöbb kromoszómánkból egy pár, azaz kettő darab van jelen minden sejtünkben. Különbséget csak a szex-kromoszómák képeznek, az X és az Y. Előbbi mindkét nemben jelen van, a női sejtekben ráadásul páros formában, az Y kromoszóma azonban csak a férfiak sejtjeiben fordul elő, az ő X kromoszómájuk satnya párjaként. Így aztán az Y kromoszómák (normális esetben) soha nem fordulnak elő kettesével, aminek következtében nem lehetnek alanyai sem a többi kromoszóma genetikai anyagát meiotikus sejtosztódásokkor összekeverő rekombinációnak (lásd jobboldali ábra).
Arra talán mindenki emlékszik gimis tanulmányaiból, hogy a legtöbb kromoszómánkból egy pár, azaz kettő darab van jelen minden sejtünkben. Különbséget csak a szex-kromoszómák képeznek, az X és az Y. Előbbi mindkét nemben jelen van, a női sejtekben ráadásul páros formában, az Y kromoszóma azonban csak a férfiak sejtjeiben fordul elő, az ő X kromoszómájuk satnya párjaként. Így aztán az Y kromoszómák (normális esetben) soha nem fordulnak elő kettesével, aminek következtében nem lehetnek alanyai sem a többi kromoszóma genetikai anyagát meiotikus sejtosztódásokkor összekeverő rekombinációnak (lásd jobboldali ábra).
Genetikai állományunk egyetlen más DNS darabja van csak még ennyire „kivételezett” helyzetben, ez pedig a sejtjeink energiát termelő speciális sejtszervecskéinek, a mitokondriumoknak a belsejében elhelyezkedő DNS molekula (mtDNS). Míg az Y kromoszóma csak apai ágon öröklődhet, addig az mtDNS-el épp fordított a helyzet, ez csak anyáról szállhat lányára. Éppen ennek a sajátos öröklődésnek köszönhető, hogy ez a két DNS szakasz jól alkalmazható Földön élő bármely ember apai, vagy anyai ágának feltérképezéséhez.
De miért is lényeges, hogy nem-rekombinálódó DNS-t vizsgáljunk? Azért, mert lényegesen leegyszerűsíti a dolgunkat, ha tudjuk, hogy a adott DNS darabok kvázi háborítatlanul kerülnek generációból, generációba, és az új változások megjelenésének és rögzülésének egyetlen módja, ha egy leendő szülő ivarsejtjeiben valami okból kifolyólag egy mutáció következik be. Így aztán, ismerve a mutációk megjelenésének gyakoriságát, a már említett rekombináció hiányában sokkal könnyebb a egyes mutációk megjelenésének idejét és helyét megsaccolni.
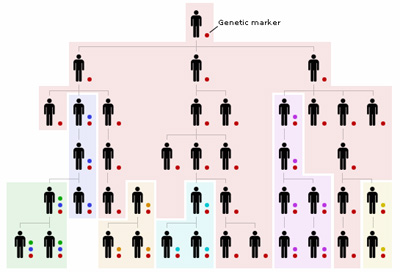 A mellékelt ábrán látható, hogy kb miként is működik mindez. A kezdeti ős összes leszármazottja hordozni fogja a kezdeti DNS mutációt (amit ezután genetikai markerként aposztrofálunk), mintegy igazolva a rokonságukat. De a második illetve a negyedik generációban újabb markerek jelennek meg, amelyek kisebb csoportok rokonságát tanusíthatják. Egyes markerek egyedi kombinációját nevezzük haplocsoportnak és minnél több markert használunk, annál szorosabb rokonsági-kötelék létét lehet megállapítani (vagy éppen a rokonság hiányát, hiszen egyes markerek nemmegléte ugyanúgy árulkodó lehet).
A mellékelt ábrán látható, hogy kb miként is működik mindez. A kezdeti ős összes leszármazottja hordozni fogja a kezdeti DNS mutációt (amit ezután genetikai markerként aposztrofálunk), mintegy igazolva a rokonságukat. De a második illetve a negyedik generációban újabb markerek jelennek meg, amelyek kisebb csoportok rokonságát tanusíthatják. Egyes markerek egyedi kombinációját nevezzük haplocsoportnak és minnél több markert használunk, annál szorosabb rokonsági-kötelék létét lehet megállapítani (vagy éppen a rokonság hiányát, hiszen egyes markerek nemmegléte ugyanúgy árulkodó lehet).
Ennek az elvnek az egyik izgalmas alkalmazása, hogy megkeressük minden ma létező mtDNS és Y kromoszóma ősét, pontosabban azok hordozóit, vagyis azt a nőt és férfit, aki minden élő ember (egyik) őse lehet. Őket a populáris irodalom „mitokondriális Évának” és „Y kromoszómális Ádámnak” nevezi, ami kétségtelenül hangzatos név, csak, ha nem vagyunk tisztában a jelentésével, könnyen félreértésekre adhat okot. Tehát mi az amit nem jelent: ez az Éva és ez az Ádám nem az első emberpár, nem az első két Homo sapiens. Nem nekik volt az első emberi mtDNS-ük/Y kromoszómájuk, ha belegondolunk egyszerűen ők azért vannak kivételezett helyzetben, mert az összes kortárs nőnek később megszakadt a lány-utódlása (pl. csak fia született, akiből aztán már nem tudott tovább adódni a mtDNS), és valami hasonló következett be a férfiakkal is fiú utód témakörben. „Mitokondriális Éva” és „Y kromoszómális Ádám” minden valószínűség szerint nem is ismerte egymást, hiszen Éva kb. 170,000 éve, Ádám pedig csak mintegy 60-75,000 éve (s bár ezek a dátumok az egyre nagyobb mintaszámok miatt még egy kicsit változhatnak, nem tűnik esélyesnek, hogy fedésbe kerülnek). (A különbség oka arra vezethető vissza, hogy a férfiak a történelem hajnalán előszeretettel választották a „háremes” életformát, s így míg egy-egy csoportban a rangidős férfinak számos gyereke született, addig a hierarchia alsó szintjein dekkolóknak csak elvétve volt esélye a szaporodásra. Így aztán egy-egy sikeres és hosszú életű férfi, viszonylag rövid idő alatt egy-egy populáció „Y kromoszómális Ádámjává” válhatott – miközben a mtDNS öröklődése jóval egalitáriusabban történt.)
Szóval akkor megvan a történet kezdőpontja: mintegy 60,000 éve valahol Ny-Afrikában már létezett egy törzs, amelyik már fellelhető volt minden ma élő ember mtDNS-ének és minden ma élő férfi Y kromoszómájának őse is. Ez a csoport, illetve ennek a csoportnak aztán egy (vagy több, lásd alább) szelete elhagyta Afrikát, hogy a többi kontinenst is meghódítsa, az ottmaradók pedig az őshaza belakásával lettek elfoglalva.
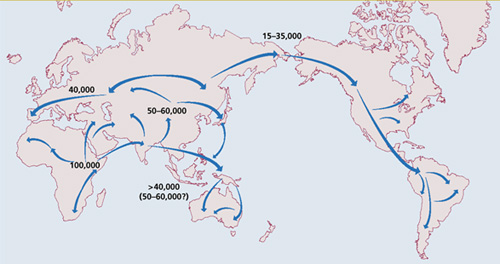
Az első nagy migrációs hullám az Indiai Óceán partjai mentén haladt és bizony messzire jutott: egész pontosan Ausztráliáig – ami azért akkor még könnyebb volt, mert az éppen tomboló jégkorszak következtében a tengerszint mintegy 100 méterrel alacsonyabban volt, így az Austrália északi partjain meghúzódó apró szigetek, még összefüggő földrészt alkottak, majdnem teljes átjárást biztosítva DK-Ázsiába. Ennek a vándorlásnak az emlékét őrzi az ún. „C haplocsoport” az Y kromoszómán és az „M haplocsoport” az mtDNS-ben, amelyek az ausztrál bennszülöttek domináns haplocsoportjai, de máshol a világon szinte sehol nem fordulnak elő, csak néhány indiai, part mentén élő közösségben.
Nem teljesen világos, hogy ezeknek az első pioníroknak egy csoportja elidőzött egy kicsit valahol Közép-Keleten és onnan indították úgy 40-50,000 éve második terjeszkedési hullámukat, vagy ez utóbbiért, a pionírok nyomán Afrikából érkező újabb csoport tehető felelőssé. Bárhogy is volt, hamarosan már emberek taposták Közép Ázsia földjét, hogy innen indulva, a földrajzi lehetőségeket kihasználva meghódítsák mind Dél-Kelet és Kelet Ázsiát, mind pedig Európát. Előbbi terjeszkedés aztán ismét csak egy nagyobb vándorlás kezdetének bizonyult (bár önmaga sem volt kicsinek nevezhető). Ezek az emberek egyre jobban képesek lettek a jeges északi tájakon is megvetni a lábukat, majd végül az így szerzett tapazstalatoknak köszönhetően, durván 20,000 éve egy ősi törzs sikeresen átkelt a Bering-szoroson, és letelepedett Alaszkában. Az Amerikai kontinens meghódításával még várniuk kellett, mert a jégkorszak, ami lehetővé tette, hogy száraz lábbal átkeljenek Ázsiából É-Amerikába, egyszersmind vastag jégtakaró alá vonta Kanada nagy részét, elvágva az utat a déli terjeszkedés elől. Arra csak a jégkorszak végén kerülhetett sor, de végül megtörtént s így mindkét amerikai kontinens bennszülött lakossága az Y kromoszomális „Q haplocsoportba” tartozik. (Az igazság az, hogy vannak – vitatott – régészeti leletek, amelyek egyesek szerint D-Amerikában már 50,000 éve ember jelenlétére utalnak, hogy ezek a leletek valósak-e vagy sem, illetve, hogy a Homo genus melyik tagja hagyhatta maga után az még nem eldöntött vita, nem is mennék bele.)
Az Európába úgy 40,000 éve megérkező H. sapiens-t egy távoli rokona várta, a neandervölgyi ember. A találkozás nem volt valami viharos (a közhidelemmel ellentétben arra semmi bizonyítk nincs, hogy a gonosz cro-magnoniak lemészárolták a szelíd nenadervölgyieket), de a két Homo genus azonos nichet töltött be, ami óhatatlanul is versenyhelyzetet teremtett, s ebből mi kerültünk ki győztesen. Persze ez a 40,000 évvel ezelőtti népvándorlás koránt sem az utolsó volt fajunk történetében, az „öreg kontinenst” becélzók közül, és még jópárral találkozhatunk a későbbi évezredekben, a legfontosabbal úgy 10,000 évvel ezelőtt, a mezőgazdaság elterjedésével tandemben, azzal szorosan összefüggve. De ez már egy másik történet.
(Bővebben még lásd a NatGeo Genographic Projectjének honlapján illetve Raskó István előadása a Mindentudás Egyetemén.)
Spencer Wells (2006) Deep Ancestry (ISBN 0-7922-6215-8)
Luigi Luca Cavalli-Sforza (2001) Genes, Peoples and Languages (ISBN 0-14-029602-6) (magyarul: Genetikai átjáró)
Behar DM, Rosset S, Blue-Smith J, Balanovsky O, Tzur S, et al. (2007) The Genographic Project Public Participation Mitochondrial DNA Database. PLoS Genet 3(6): e104 doi:10.1371/journal.pgen.0030104
Williamson SH, Hubisz MJ, Clark AG, Payseur BA, Bustamante CD, et al. (2007) Localizing Recent Adaptive Evolution in the Human Genome. PLoS Genet 3(6): e90 doi:10.1371/journal.pgen.0030090
Shriver MD, Kittles RA (2004) Genetic ancestry and the serch for personalized genetic histories. Nat Rev Gen 5: 611-618.
Bamshad M, Wooding S, Salisbury BA, Stephens JC (2004) Deconstructing the relationship between genetics and race. Nat Rev Gen 5: 598-609.
Cavalli-Sforza LL, Feldman MW (2003) The application of molecular genetic approaches to the study of human evolution. Nat Gen 33: 266-275.
Egy Spencer Wells is megjelent a Talentumnál Az ember útja címen.
http://bookline.hu/control/producthome?id=36847&type=22
Két kérdésem van:
1. Ezek a vizsgálatok egyértelműen kizárták az indiánok legalább részleges polinéziai származását?
2. Itt 100 000 évvel ezelőttben adjátok meg a terjeszkedés kezdetét. (Olvastam állítólag ennél régebbi közelkeleti leletekről is, de most nem ez a lényeg.) Ezek szerint az emberiség még ez után is sokáig egységes populációt alkotott?
(Hisz másképp nem lehetne 30 000 évvel későbbi az Y kromoszomális Ádám.)
Még egy magánvélemény a Neandervölgyiekhez. Azt mondjátok semmi bizonyíték arra, hogy kiírtotta őket a sapiens. O.K. Nyilván nem volt szervezett írtás vagy akár folyamatos háború, de ha két lényegében azonos életformát folyatató vadászó faj egy területen él, nehéz elképzelni, hogy ne lennének időnként összecsapások. (A sapiens törzseken belül is voltak/vannak.) És ha vannak, akkor azok kimetele meghatározó lehet abból a szempontból, hogy ki szorít ki kit.
Ő, elnézést én úgy tudtam, hogy a homo sapiens kialakulását 100 000 évre teszik. Rosszul tudom?
Mackosajt,
a teljes leszarmazast igen. De, hogy kisebb merteku keveredes volt-e, arrol nem mondanak semmit (lasd kovetkezo bejegyzes).
Mackosajt&Mikrobi,
az anatomiailag modern ember kialakulast 100-200,000 eve teszik (erzesem szerint a konszenzus utobbihoz van kozelebb, mostansag). Azaz eleg sok bizonytalansag van a pontos datum korul – akarcsak az altalam idezett Evas es Adamos datumok eseteben. Amiket irtam, ma azokat tartjuk a legvaloszinubbnek, de minden esetben valojaban van egy tobb ezer (neha tizezer) eves hibahatar. Miutan kialakult a modern ember hosszabb ideig valszleg viszonylag kis teruleten elt, ha nem is egy populacioban, de mindenkeppen egymassal rendszeres kontaktusban levo populaciokban. (Persze az is lehet, hogy tobb izolalt populacio volt, de csak az egyik maradt fenn&terjeszkedett sikeresen. Oszinten, nem tudom, hogyan lehetne kulonbseget tenni a ket lehetoseg kozott, a mai adatokkal. De en nem vagyok populaciogenetikus, igy ez nem jelent semmit :-).)
Az Ádám-Éva elnevezések amennyire hangzatosak, annyira félrevezetőek is. Valójában ne egy ősapát és ősanyát kell elképzelni, hanem azt, hogy (legalább) két alkalommal a sapiens története során volt egy-egy úgy nevezett üvegnyak (bottle neck), amikor nagyon lecsökkent a populációméret. Legalábbis annak a populációnak a mérete, amiből a későbbi sapiensek lettek. Nincs feltétlenül szó egy nőről és egy férfiról csak arról, hogy olyan kicsi lett a változatosság, ami akár egy-egy személyt is jelenthet. Kis populációkban ráadásul pusztán a véletlen miatt is eltűnhetnek változatok.
Jól ismert példa erre a Bounty lázadóinak esete, akik a Pitcairn-szigeten telepedtek le. 200 év alatt az angol vezetéknevek fele eltűnt, pusztán csak a véletlen miatt (nem volt gyereke vagy csak lánya született a név viselőjének). Könnyen lehet, hogy ehhez hasonlóan maradt meg Ádám Y kromoszómája és Éva mitokondriuma is.
Egyes feltételezések szerint Éva esetében a sapiens kialakulása volt az, ami olyan apró populációban játszódott le, Ádámnál pedig az utolsó nagy eljegesedés miatt fogyatkozott meg a sapiens populáció.
Az lenne a kérdésem, hogy,
van-e bizonyíték arra, hogy az L3 mtDNSből feljődött ki az N és M mtDNS és ha van, akkor mi az?
sinus,
szerintem itt biztos megvan amit keresel:
http://www.phylotree.org/tree/main.htm