
 A többsejtűség eredete a földi élet egyik fordulópontja lehetett, nem véletlen, hogy ma is sokan igyekeznek megfejteni, hogy miképpen is történhetett. Az, hogy feltehetőleg többször is bekövetkezett ez az átmenet, elvileg könnyebbé teheti a mögötte álló mechanizmus feltérképezését, nem véletlen, hogy sokan érdeklődnek a téma iránt.
A többsejtűség eredete a földi élet egyik fordulópontja lehetett, nem véletlen, hogy ma is sokan igyekeznek megfejteni, hogy miképpen is történhetett. Az, hogy feltehetőleg többször is bekövetkezett ez az átmenet, elvileg könnyebbé teheti a mögötte álló mechanizmus feltérképezését, nem véletlen, hogy sokan érdeklődnek a téma iránt.
Sexcomb kolléga a Chlorellákon végzett kísérleteket mutatta be nem is olyan régen, én pedig pár éve a Volvox-ok apropóján írtam arról, hogy mit is gondolunk a többsejtű életformák kialakulásáról.
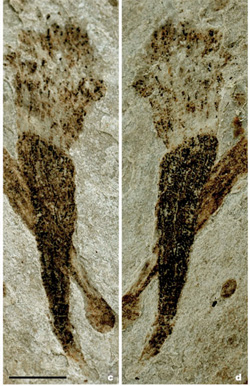
Bármi volt is a valódi molekuláris mechanizmus, biztos, hogy megtörtént és az ediakara időszakban, durván 570 millió éve, nem is olyan sokkal a „hógolyó Földként” aposztrofált globális eljegesedések után, az óceánok mélyét már az ekkortájt megjelenő, komplex többsejtű szervezetek uralták. (Erről az időszakról (is) szól egyébként Attenborough legutóbbi filmsorozata, aki teheti, nézze meg. Reklámblokkunk olvasták. ;-))
Most épp Kínából került elő számos, kifejezetten érdekes fosszília, amelyek egy ún. Lantian bióta képviselői lehettek. A Lantian az eddig ismert legősibb ediakarai együttesnél, az Avalonnál is idősebb lehet a becslések szerint, és nagyon másképpen is néz ki. Ami összességében azt sugallja, hogy az élet már ebben az egészen ősi korban is igencsak sokszínű és gyorsan változó lehetett.

Yuan X, Chen Z, Xiao S, Zhou C, Hua H. (2011) An early Ediacaran assemblage of macroscopic and morphologically differentiated eukaryotes. Nature 470: 390-393.
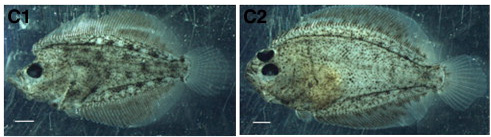 Pár évvel ezelőtt
Pár évvel ezelőtt 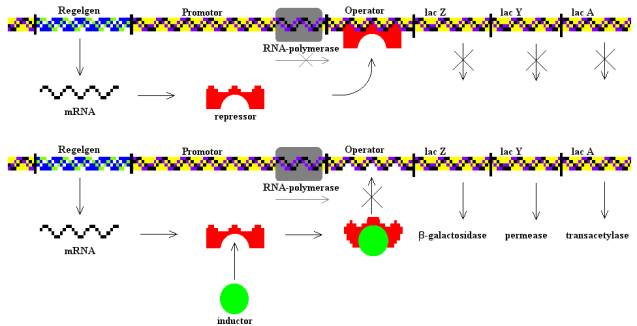
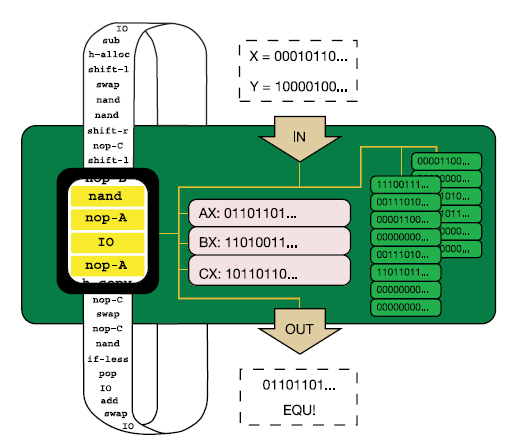 Richard E. Lenski és munkatársai éppen erre vállalkoztak, számítógépes programokkal, mondhatni digitális élőlényekkel modellezték egy bonyolult működés kialakulását. Mint tudjuk, az evolúció három alapfeltétele, hogy a résztvevő egységek önmagukat lemásolják, szaporodjanak (1), magukhoz hasonló utódokat hozzanak létre, hogy ezekben a másolatokban időnként hibák, mutációk történjenek (2) és végül, hogy az egyes változatok életképessége különbözzön, (3) azaz szelekciós nyomás hasson az egységekre. A kísérletben használt digitális élőlények ezt a három feltételt teljesítették, önmagukat másolták, időnként hibásan és végig versengtek egymással. A modellhez az
Richard E. Lenski és munkatársai éppen erre vállalkoztak, számítógépes programokkal, mondhatni digitális élőlényekkel modellezték egy bonyolult működés kialakulását. Mint tudjuk, az evolúció három alapfeltétele, hogy a résztvevő egységek önmagukat lemásolják, szaporodjanak (1), magukhoz hasonló utódokat hozzanak létre, hogy ezekben a másolatokban időnként hibák, mutációk történjenek (2) és végül, hogy az egyes változatok életképessége különbözzön, (3) azaz szelekciós nyomás hasson az egységekre. A kísérletben használt digitális élőlények ezt a három feltételt teljesítették, önmagukat másolták, időnként hibásan és végig versengtek egymással. A modellhez az 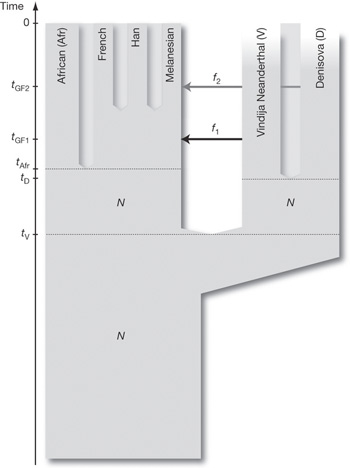 Az idei év egyik nagy dobása kétségtelen az archaikus Homo fajok genomjainak publikálása lett. Azért használok többesszámot, mert a májusi
Az idei év egyik nagy dobása kétségtelen az archaikus Homo fajok genomjainak publikálása lett. Azért használok többesszámot, mert a májusi  A csigaház csavarodik, ez etalon. De a csavarodás iránya, az iránynak a miértje illetve a csavarodás irányának megváltozása már koránt sem az.
A csigaház csavarodik, ez etalon. De a csavarodás iránya, az iránynak a miértje illetve a csavarodás irányának megváltozása már koránt sem az.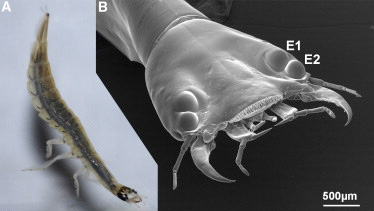 A szem evolúciója hálás téma, a különleges szemeké pedig duplán az. Írtunk már ennek apropóján
A szem evolúciója hálás téma, a különleges szemeké pedig duplán az. Írtunk már ennek apropóján  Eukarióták közti horizontális gén-transzfer (HGT) mindig izgalmas téma, mert ritka, amikor megtörténik az általában különösen érdekes (mármint, hogy milyen hasznot hoz az új gazdájának a beépült gén), és nem utolsó sorban azért is érdekes, mert ez a GMO-ellenes tábor nagy zsákosembere, amitől félni kell.
Eukarióták közti horizontális gén-transzfer (HGT) mindig izgalmas téma, mert ritka, amikor megtörténik az általában különösen érdekes (mármint, hogy milyen hasznot hoz az új gazdájának a beépült gén), és nem utolsó sorban azért is érdekes, mert ez a GMO-ellenes tábor nagy zsákosembere, amitől félni kell. Lassan több mint száz éve, hogy
Lassan több mint száz éve, hogy  Amikor két napja megegyeztünk a
Amikor két napja megegyeztünk a