 A kreacionisták egy szűnni nem akaró tévedése/csúsztatása az evolúcióval kapcsolatban, hogy a folyamatnak teljesen véletlenszerű a kimenetele. Ez persze ebben a formában merőben hamis, hiszen maga a természetes szelekció egy nagyon is rigorózus válogatás során „dönti el”, hogy adott faj mely egyedei talpraesettebbek adott körülmények között, ugyanakkor annyi igazságmag azonban van benne, hogy magának az evolúciós mechanizmusnak létezik egy véletlen komponense is. Ez pedig nem más, mint azoknak a változatoknak a létrejötte, amelyekből a szelekció később „válogat”.
A kreacionisták egy szűnni nem akaró tévedése/csúsztatása az evolúcióval kapcsolatban, hogy a folyamatnak teljesen véletlenszerű a kimenetele. Ez persze ebben a formában merőben hamis, hiszen maga a természetes szelekció egy nagyon is rigorózus válogatás során „dönti el”, hogy adott faj mely egyedei talpraesettebbek adott körülmények között, ugyanakkor annyi igazságmag azonban van benne, hogy magának az evolúciós mechanizmusnak létezik egy véletlen komponense is. Ez pedig nem más, mint azoknak a változatoknak a létrejötte, amelyekből a szelekció később „válogat”.
Ezek a változatok mutációk során keletkeznek (amely minden osztódó sejt sajátossága) és hogy épp melyik kromoszóma, melyik bázispárját érintik kvázi véletlenszerű (valójában nem teljesen, de az egy hosszú, külön sztori). Vagyis, ha, Stephen Jay Gould sokat vitatott hasonlatával élve, visszapörgetnénk az élet filmjét egy korábbi időpontra és újból forogni engednénk, akár teljesen más kimenetele is lehetne a mozinak, mint amit mi ismerünk. (Akár, de mint sokan hangsúlyozzák, nem feltétlenül.)
Természetesen globális „rewind” gomb hiányában mindez gondolatkísérlet marad, viszont ha hajlandóak vagyunk kicsiben gondolkodni, tervezhetünk olyan kísérletet, ami valami hasonlót tesztel. Az alapfelállás egyszerű: egy baktérium (legyen mondjuk a legkedveltebb, az Escherichia coli) tenyészetét tucatnyi kémcsőbe töltjük szét, majd a kémcsövek tartalmának egy kis részét minden reggel új kémcsőbe és tápoldatba löttyintjük. Közben minden x. generációt lefagyazstjuk, hogy ha akarjuk onnan újból futtathassuk a kísérletet. Mivel, mint az előbb már említettem, a bacik osztódásaik során akarva-akaratlanul mutációkat szednek össze, amelyeket a kémcsőben (is) zajló természetes szelekció gyorsan „elbírál”, ha elég sokáig csináljuk a dolgot, végül az aktuális tucatnyi kémcsövünk mindegyikében egy-egy olyan baktérium generáció fog éldegélni, amely nemcsak a kezdeti őstől különbözik sokban, de a másik tizenegy kémcső mindegyikétől is.
Richard Lenski laborjában pont egy ilyen kísérlet zajlik már vagy 20 éve, azzal az extra kis csavarral ellátva, hogy a tápoldat csak épp minimális mennyiségben tartalmazza a bacik által igencsak kedvelt cukrot, a glükózt, viszont sok van benne egy másik vegyületből, citrátból, amelyet viszont standard körülmények között képtelenek felvenni. Mivel a sejt belső mechanizmusai már lehetővé tennék, hogy hasznosítsa a citrátot, ha valahogy egy baktérium „megoldaná”, a molekula átmozgatását a sejtfalán és membránján, olyan előnyre tenne szert, amivel kiemelkedne társai közül (a kiemelkedés természetesen gyors elszaporodásban nyilvánulna meg). Ez egyszerűen hangzik, mégis sok évnek és közel 33 000 bakteriális generációnak kellett eltelnie ahhoz, hogy a tizenkét kémcső egyikében ez megtörténjen. Vagyis vagy valami nagyon-nagyon ritka és egzotikus egyszeri mutáció okozta, vagy a sok ezer generáció alatt felhalmozódott más mutációk tették végül lehetőve, hogy megjelenjen az az egy is, amelyik a hatékony citrát használatot valósággá váltotta.
Az ötszáz generációnként lefagyasztott minták révén Lenskiéknek alkalmuk volt tesztelni, mi is a helyzet. Először is azt nyomozták ki, hogy valahol a 31 000. és 31 500. generáció közt jelent meg a citrát hasznosítás a szóbanforgó kémcső-populációban (nevezzük az egyszerűség kedvéért ezután ezt a jelleget „Cit+„-nak). Ezek a korai bacik azonban még nem voltak túl hatékonyak az új élelmiszerforrás hasznosításában, így nem sokkal később majdnem kiszorította őket a glükózt egyre hatékonyabban hasznosító társaik, Végül egy újabb mutáció eredményeként a citrát felhasználás hatékonysága lényegesen javult, s a Cit+ sejtek közel egyeduralkodóvá nem váltak ebben a kémcsőben.
Ez azonban még nem a teljes történet, Lenskiéknek az is feltűnt hogy az „újrafuttatott szalagok” esetében , több ismétlés mellett is csak ezen az egy vonalon, és mindig csak a 20 000. generáció után induló vonalakból alakultak ki Cit+ sejtek. Azaz valamikor ekkortájt honosodott meg egy olyan mutáció ebben a kémcsőben, amely lehetővé tette, hogy később a citrát-evés kialakulhasson (bár önmagában még kevés lett volna ahhoz). Ha véletlenül nem alakult volna ki (mint ahogy a 20 000. generációnal korábbi mintákból induló párhuzamos kísérletek többségében nem is jelent meg), akkor ezen „történeti örökség” hiányában nem jöhetett volna létre a Cit+ fenotípus. (Még fontos hangsúlyozni, hogy ez az első mutáció vagy semleges volt, vagy esetleg már akkor és ott volt valami közvetlen – a citráttól tök független – előnye, hiszen az evolúció nem tervez előre.)
A nagy kérdés persze az ezekután, hogy mi is ez a három mutáció, milyen géneket érintenek, stb. Erre azonban várni kell, bár kétségtelen, hogy Lenski és társai már gőzerővel kutatják ezt.
(A téma iránt jobban érdeklődőknek ajánlott olvasmány még PZ Myers postja is, amelyben Michael Behe kötekedését szedi atomjaira. A kötött bacik fényképe a Craftzine.com blogról származik.)
Blount ZD, Borland CZ, Lenski RE (2008) Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. PNAS 105(23): 7899-7906.

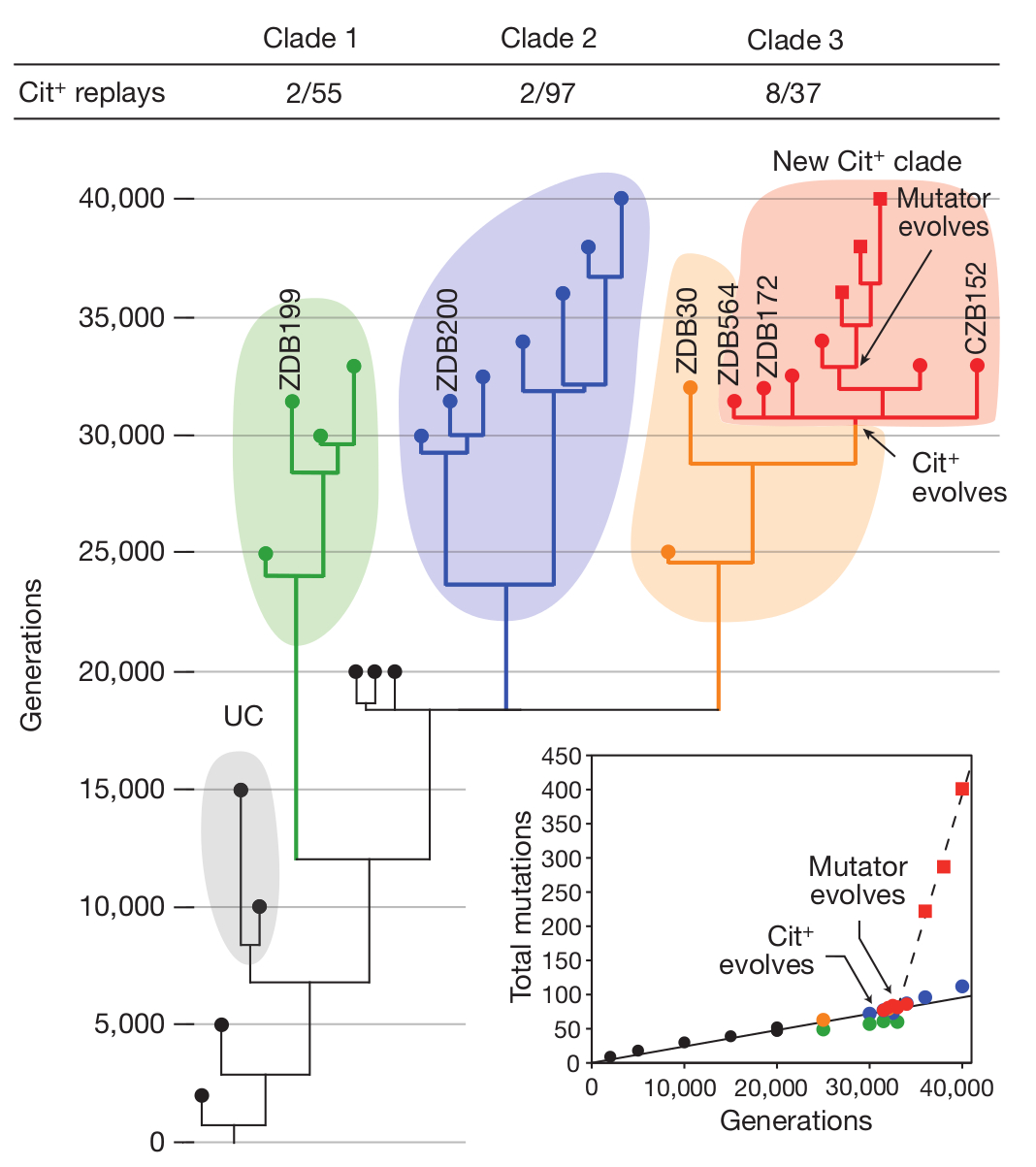 Richard Lenskit nyilván senkinek sem kell bemutatnom, lebilincselően izgalmas kísérletei (1, 2, 3) elég sok adalékot szolgáltattak az evolúció megismeréséhez. Kétségkívül leghíresebb munkája az évtizedek óta folyó E. coli evolúciós kísérlete, ahol egy E. coli törzset többfelé osztott, majd szőlőcukorban szegény, citromsavban gazdag táptalajra helyezte őket és figyelte, mi történik velük. Az E. coli egyik jellemzője ugyanis, hogy csak anaerob körülmények között képes citromsavat hasznosítani, oxigén jelenlétében a citromsav nem jut át a sejtmembránon, így nem is képes táplálékként felhasználni a sejt. A mikrobiológiai gyakorlatban ez alapján azonosítják E. coliként. Lenski eredeti feltételezése az volt, hogy mivel az E. coli anaerob körülmények között képe citromsavat is hasznosítani, megfelelő körülmények között előbb-utóbb az evolúció során kialakul az oxigén jelenlétében való citromsavfogyasztás képessége is. Nem csalódott, röpke 31000 nemzedék után a tizenkét törzs egyikében meg is jelentek citromsavat is hasznosító sejtek, ezeket nevezte el Cit+ -nak.
Richard Lenskit nyilván senkinek sem kell bemutatnom, lebilincselően izgalmas kísérletei (1, 2, 3) elég sok adalékot szolgáltattak az evolúció megismeréséhez. Kétségkívül leghíresebb munkája az évtizedek óta folyó E. coli evolúciós kísérlete, ahol egy E. coli törzset többfelé osztott, majd szőlőcukorban szegény, citromsavban gazdag táptalajra helyezte őket és figyelte, mi történik velük. Az E. coli egyik jellemzője ugyanis, hogy csak anaerob körülmények között képes citromsavat hasznosítani, oxigén jelenlétében a citromsav nem jut át a sejtmembránon, így nem is képes táplálékként felhasználni a sejt. A mikrobiológiai gyakorlatban ez alapján azonosítják E. coliként. Lenski eredeti feltételezése az volt, hogy mivel az E. coli anaerob körülmények között képe citromsavat is hasznosítani, megfelelő körülmények között előbb-utóbb az evolúció során kialakul az oxigén jelenlétében való citromsavfogyasztás képessége is. Nem csalódott, röpke 31000 nemzedék után a tizenkét törzs egyikében meg is jelentek citromsavat is hasznosító sejtek, ezeket nevezte el Cit+ -nak.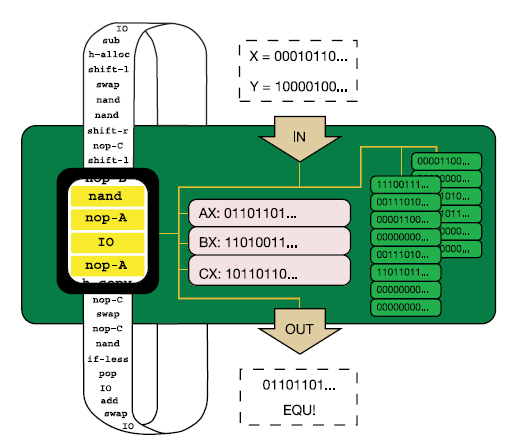 Richard E. Lenski és munkatársai éppen erre vállalkoztak, számítógépes programokkal, mondhatni digitális élőlényekkel modellezték egy bonyolult működés kialakulását. Mint tudjuk, az evolúció három alapfeltétele, hogy a résztvevő egységek önmagukat lemásolják, szaporodjanak (1), magukhoz hasonló utódokat hozzanak létre, hogy ezekben a másolatokban időnként hibák, mutációk történjenek (2) és végül, hogy az egyes változatok életképessége különbözzön, (3) azaz szelekciós nyomás hasson az egységekre. A kísérletben használt digitális élőlények ezt a három feltételt teljesítették, önmagukat másolták, időnként hibásan és végig versengtek egymással. A modellhez az
Richard E. Lenski és munkatársai éppen erre vállalkoztak, számítógépes programokkal, mondhatni digitális élőlényekkel modellezték egy bonyolult működés kialakulását. Mint tudjuk, az evolúció három alapfeltétele, hogy a résztvevő egységek önmagukat lemásolják, szaporodjanak (1), magukhoz hasonló utódokat hozzanak létre, hogy ezekben a másolatokban időnként hibák, mutációk történjenek (2) és végül, hogy az egyes változatok életképessége különbözzön, (3) azaz szelekciós nyomás hasson az egységekre. A kísérletben használt digitális élőlények ezt a három feltételt teljesítették, önmagukat másolták, időnként hibásan és végig versengtek egymással. A modellhez az