
 A csigaház csavarodik, ez etalon. De a csavarodás iránya, az iránynak a miértje illetve a csavarodás irányának megváltozása már koránt sem az.
A csigaház csavarodik, ez etalon. De a csavarodás iránya, az iránynak a miértje illetve a csavarodás irányának megváltozása már koránt sem az.
Márpedig, bár a laikus szemlélőnek szinte mindegynek tűnhet, hogy merre csavarodik egy csiga háza (esetleg még az is meglepő, hogy fajon belül többé-kevésbé fix az irány, amely genetikailag kódolt), a csigák és a velük ökölógiailag szoros kapcsolatban élők számára igen fontos a kiszámíthatóság. Ahhoz, hogy ez a kiszámíthatóság felboruljon, komoly okra van szükség, de nem lehetetlen.
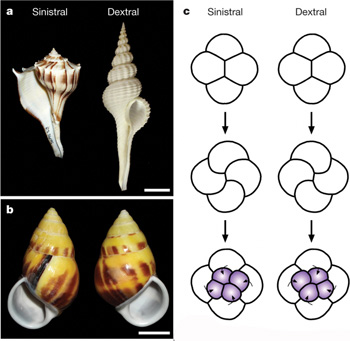 Különböző csigafajok esetében találhatunk az óramutató járása irányában csavarodó házakat (dextralis), illetve azzal ellenkező irányban kunkorodókat (sinistralis). A folyamat, ami végül is nem más mint a bal-jobb aszimmetria kialakulása egy puhatestűre levetítve, a fejlődés kezdetén lezajló osztódásokra vezethető elsődlegesen vissza, amelyek később jellegzetes génexpressziós változásokat okoznak, és ezek hozzák létre a sztereotip irányba kunkorodó házakat.
Különböző csigafajok esetében találhatunk az óramutató járása irányában csavarodó házakat (dextralis), illetve azzal ellenkező irányban kunkorodókat (sinistralis). A folyamat, ami végül is nem más mint a bal-jobb aszimmetria kialakulása egy puhatestűre levetítve, a fejlődés kezdetén lezajló osztódásokra vezethető elsődlegesen vissza, amelyek később jellegzetes génexpressziós változásokat okoznak, és ezek hozzák létre a sztereotip irányba kunkorodó házakat.
A csigaembró fejlődése az Ősszájúakra jellemző spirális osztódás szerint zajlik, ami tömören azt jelenti, hogy az egyes sejtek osztódási síkjai nem párhuzamosak, hanem adott szöget zárnak be egymással. Ennek következtében nyolcsejtes stádiumtól a fejlődő embrió különböző síkjaiban levő sejtek már nem pont egymás alatt vannak, így felülről nézve egy spirális szerveződés illúzióját keltik. Hogy milyen konkrét szögben térnek el az osztódási síkok az azonban nincs kőbe vésve, s ennek megfelelően, attól függően, hogy jobbra vagy balra történik a deviáció, alakul ki később a csigaház csavarodásának iránya.
Annyira így van ez, hogy bár a folyamat (az osztódási sík kialakulása) genetikailag szabályozott (lásd mindjárt), ha apró üvegbotok segítségével ellentartunk az első aszimmetrikus osztódásnak, és az ellenkező irányban toljuk el a sejteket, mint azt természetes módon maguktól tennék, akkor ellenkező csavarodású csigaházat kapunk! (Nagyon türelmes japán kutatók végezték el a kísérletet, ki más :-).)
Bár az embrió spiralitásából már látszólag jól le lehetne vezetni, hogy miként jön létre az adott fajra specifikus csigaház, igazából van egy nem elhanyagolható köztes lépés: néhány jellegzetes gén specifikus, aszimmetrikus kifejeződése. Ha utóbbit meggátoljuk, akkor a csigaház is elveszti a csavarodását, nem csavarodik semerre. (A történet slusszpoénja, hogy ezek a „jellegzetes gének” ugyanazok, amelyek a gerincesek aszimmetriájáért is felelősek, így valószínűleg a kétoldali-szimmetriájú közös ős mintázatképződéséért is felelhettek.)
 Visszatérve azonban a kezdeti osztódásokhoz, ezekben az osztódási sík irányát egy „anyai faktor” határozza meg, azaz egy olyan, az anya által a petesejtbe juttatott mRNS vagy fehérje, amely az osztódási folyamat szabályozója. Ha ez valami követekeztében mutálódik, és a síkok ellenkező irányba eltolódnak, vagyis egy dextrális fajban sinistralis egyedek jelennek meg, akkor kész a baj: nem lesznek képesek párzani.
Visszatérve azonban a kezdeti osztódásokhoz, ezekben az osztódási sík irányát egy „anyai faktor” határozza meg, azaz egy olyan, az anya által a petesejtbe juttatott mRNS vagy fehérje, amely az osztódási folyamat szabályozója. Ha ez valami követekeztében mutálódik, és a síkok ellenkező irányba eltolódnak, vagyis egy dextrális fajban sinistralis egyedek jelennek meg, akkor kész a baj: nem lesznek képesek párzani.
A szárazföldi csigafajok hermafroditák, de egymás kölcsönös megtermékenyítéséhez két példány megfelelő módon kell egymáshoz simulnia, hogy párzószerveik a megfelelő nyílásokba hatolhassanak (a mellékelt ábra (b) panelje). Az ellenkező irányban csavarodó házak ezt az összecsavarodást akadályozzák meg (lásd (c), ahol a fehér nyilak az összepasszíthatatlan ivarszervi nyílásokat jelzik.) Gyakorlatilag megtörtént a prezigotikus izoláció, a populáció bizonyos egyedei képtelenek egymással szapordoni, vagyis két külön fajként lehet szinte rájuk tekinteni. (Érdekes mellékvágány, egész röviden, hogy a legtöbb modell a fajképződés genetikai alapjait két lókuszhoz köti – lásd egy rövid magyarázatot itt, élesztőgombákra -, és nagyon ritka az olyan eset, ahol egyetlen gén változása speciációt eredményezhet.)
Épp ezért nagyon ritka egy csiga-populációban az ilyen irányú változás, hiszen komoly evolúciós ára van. Ugyanakkor a ritka, az nem lehetetlen. Az anyai mutációk jellegüknél fogva azt jelentik, hogy a fenotipikusan hétköznapi kinézetű, homozigóta anya összes utódja kinézetre mutáns lesz, az apától függetlenül (tehát az sem számít, hogy milyen az utódok genotípusa). Így, mivel a csigaház-csavarodás genetikai mutációját hordozó anya számára nem gond a párzás, utódai is többen lesznek (lehetnek). Azaz nem lehetetlen, hogy majd kifejlett példányként maguk is összefussanak (ha nem is gyakran) egy másik „revertált” egyeddel, és szaporodjanak. Így viszont a mutáns allél képes elvileg fennmaradni (éppen csak) a populáció genetikai össz-állományában. Bizonyos körülmények azonban előnyössé is tehetik a revertált allélt, és ilyenkor a szelekció „szárnyán” gyorsan elterjedhet.
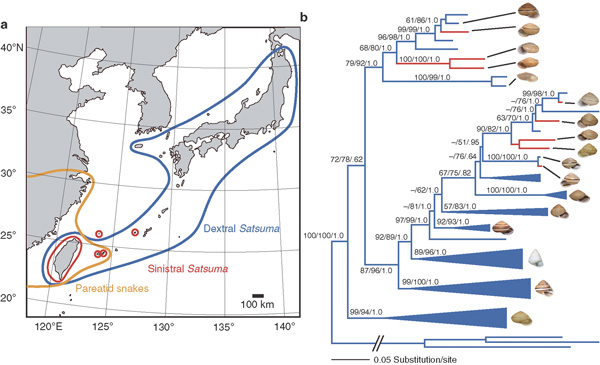
Egy, a napokban megjelent cikk (és eddig ehhez írtam lényegében a hosszas körítést) pont egy ilyen esetet mutat be. A Kelet-Ázsiában őshonos Satsuma csigafajok dextrális típusúak, de több helyen jelentős sinistrális populációk találhatóak. Az ok minden jel szerint a legfőbb ragadozójukkal folytatott evolúciós versenyben rejlik.
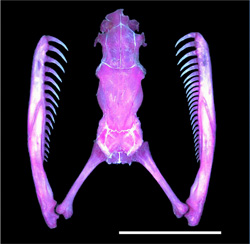 A helyi, pareatid kígyófajok ugyanis annyira specializálták magukat a döntő többségében dextrális csavarodású csigák elfogyasztására, hogy maguk is jellegzetes anatómiai, ill. viselkedésbeli aszimmetriákat fejlesztettek ki. Felső állkapcsuk egyik oldalán sokkal több fog található, és ezzel egy olyan, szekvenciális mozdulatsorozatot hajtanak végre táplálkozáskor, amelynek eredményeként a csiga a hasukban végzi. (Itt látható filmen egy tipikus vadászat.)
A helyi, pareatid kígyófajok ugyanis annyira specializálták magukat a döntő többségében dextrális csavarodású csigák elfogyasztására, hogy maguk is jellegzetes anatómiai, ill. viselkedésbeli aszimmetriákat fejlesztettek ki. Felső állkapcsuk egyik oldalán sokkal több fog található, és ezzel egy olyan, szekvenciális mozdulatsorozatot hajtanak végre táplálkozáskor, amelynek eredményeként a csiga a hasukban végzi. (Itt látható filmen egy tipikus vadászat.)
Ugyanakkor, az evolúciós alkalmazkodás, ami hatékonyabbá teszi a dextrális csigák fogyasztását, szinte elleheteteníti a sinistrális zsákmány megszerzését: a kígyó rossz oldalról próbálja megragadni a csigát, és a különleges rágómzogása is épp az ellenkező hatást éri el, vagyis a csiga kiszabadul a szájából. (Egy sikertelen vadászat videója itt található.) A csigák szempontjából a védekezés szinte tökéletes: míg egy kontroll-kísérletben az összes dextrális csigát levadászták a kígyók, addig a sinsitrális társaik 87%-a elmenekült.
Azaz ebben a közegben igen nagy evolúciós előnyt jelent egy-egy revertációt okozó mutáció. Így talán nem is meglepő, hogy a vizsgálatok szerint az egyes revertált populációk egymástól függetlenül alakultak ki, azaz többször meg tudott jelenni és el tudott terjedni ilyen hatású mutáció (ugyanakkor, hogy konkrétan, DNS szinten, milyen mutációjáról van szó, azt még nem tudjuk, így az is lehet, hogy több génről van szó, amelyek ugyanazt a sejtbiológiai folyamatot, az osztódási sík helyzetét képesek befolyásolni).
Bónusz videók és információk a kutatást vezető Masaki Hoso honlapján olvashatók.
Hoso M, Kameda Y, Wu SP, Asami T, Kato M, Hori M. (2010) A speciation gene for left-right reversal in snails results in anti-predator adaptation. Nat Commun 1(9): 133.
Kuroda R, Endo B, Abe M, Shimizu M. (2009) Chiral blastomere arrangement dictates zygotic left-right asymmetry pathway in snails. Nature 462: 790-794.
Grande C, Patel NH. (2009) Nodal signalling is involved in left-right asymmetry in snails. Nature 457: 1007-11.
Ueshima R, Asami T (2003) Evolution: single-gene speciation by left-right reversal. Nature 425: 679.
hol lehet olyan ábrákat találni mint ami a jobb alsó sarokban van, csak emlősökről?
Ha a filogenetikai fára gondolsz, akkor milyen csoportok legyenek rajta?
Az ószájúak helyesen ősszájúak lesz.
lájk, lájk… 🙂
@Syp: ez megmagyarázza, hogy miért nem talált a Google semmit tegnap ;-).
Ez állati jó! két dologban pontositanék:
– nem minden szárazföldi csiga hermafrodita csak a tüdősök
– az ellenkezö irányu csavarodás egy populáción belül nem akadálya a párzásnak csak eléggé megneheziti. A ház alakja (orsó vagy lapos) is számit a párosodási valószinüség alakulásában. Persze ezt is japánok vizsgálták 🙂
Köszi, a tüdős dolgot nem tudtam, a másikat igen, csak nem akartam tovább komplikálni a sztorit. Itt konkrétan lapos, relatíve kisméretű fajokról van szó.
Felteszem filogenetikai fára gondolok.
Emlősök jók lennének, de az igazi valami interaktív cucc lenne, beraknám a zebrát meg a réti héját, aztán megrajzolna mindent fel a közös ősig, meg onnan le minden leszármazottig.
Gondolom túl sokat akarok…
„Így, mivel a csigaház-csavarodás genetikai mutációját hordozó anya számára nem gond a párzás, utódai is többen lesznek (lehetnek).Azaz nem lehetetlen, hogy majd kifejlett példányként maguk is összefussanak (ha nem is gyakran) egy másik „revertált” egyeddel, és szaporodjanak.”
Jó értem, hogy ez azt jelenti, hogy egy ilyen „revretált” populáció alapját mindig testvérek és féltestvérek képezik? Ha igen, vajon mennyire káros ez?
Szuper a cikk (Nem kötekedésből, de a spirális barázdálódás az ősszájuakon belül csak a Lophotrochozoakra jellemző, szerintem)
Tudtok valami jo linket, ahol ami Lamarck-i evolucio cafolataval foglalkozik? Egyreszt, hogy nincs olyan mechanizmus, hogy a szulok szerzett tulajdonsagai atoroklodnek az ivarsejteken keresztul az utodnakba. Masreszt, hogy statisztikailag sem mutathato ki ilyesmi? (Egy vallas/evolcio vitaban lenne ra szuksegem)
@Wladi: itt nüanszokról van szó, szóval jó lenne tudni mi a pontos kérdés. Mert vannak az epigenetikus módosítások, amelyek laikus számára lamarcki öröklődést biztosítanak. Pl ezért van, hogy elhízott (pontosabban szándékosan felhízlalt) patkányok utódai hajlamosabbak a cukorbetegségre. De persze ez nem úgy működik, ahogy Lamarck elképzelte, mert nem az örökítőanyagban lesz a módosítás, illletve csak bizonyos határok között lehet módosítani a fejlődést.
Egyébként ez jó alapozásnak tűnik:
http://skepticwiki.org/index.php/Lamarckism
Ha már kívánságműsor van, akkor a Centriolákról nincs valami jó összefoglalótok? Rágja a fülem egy Értem rajongó, hogy azt mennyire jól magyarázza az ID…
😀
Dolphin, ez erdekes tema, irnal valamikor ezekrol az epigenetikus modositasokrol reszletesebben?