
 És ha már a közelmúltban különleges halakról esett szó, álljon itt még egy oszlopos és uszonyos példája a természetes szelekció jellegformáló erejének.
És ha már a közelmúltban különleges halakról esett szó, álljon itt még egy oszlopos és uszonyos példája a természetes szelekció jellegformáló erejének.
A levegőben és a vízben való látás kicsit különböző adaptációt igényel. A két közeg törésmutatója is eltérő, ráadásul a vízben a szem kevésbé van kitéve az UV sugárzás romboló erejének, így kevésbé kell védekeznie ellene. Nehéz helyzetben van hát az olyan (gerinces) élőlény, aki egy kicsit mindkét közegben szeretne otthonosan mozogni, hiszen azt gondolnánk, hogy vagy két extra szemet lesz kénytelen „növeszteni”, hogy a másik közegben is jól lásson, vagy valamilyen módon választania kell, melyik közegben akar jobban látni.
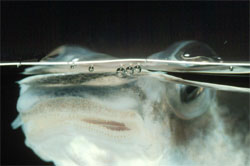
 De ha így gondolnánk, rosszul gondolnánk, mert van példa arra, hogy miképpen lehet mindkét világot gond nélkül szemmel tartani. Ez a példa pedig egy közvetlenül a vízfelszín alatt élő, kis dél-amerikai hal, az Anableps anableps, azaz helyi zsargonban „cuatro ojos”, a négyszemű hal.
De ha így gondolnánk, rosszul gondolnánk, mert van példa arra, hogy miképpen lehet mindkét világot gond nélkül szemmel tartani. Ez a példa pedig egy közvetlenül a vízfelszín alatt élő, kis dél-amerikai hal, az Anableps anableps, azaz helyi zsargonban „cuatro ojos”, a négyszemű hal.
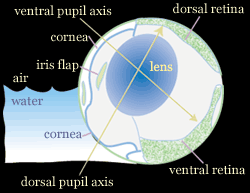
A nevével ellentétben a halnak természetesen két szeme van, de mindegyik közepén (a vízfelszín magasságában) egy kis pigment folt található, ami azt az illúziót kelti, mintha a halacskának mindkét oldalon 2-2 szeme lenne egymás felett. Az Anableps szemében két fontos adaptációra lelhetünk. Egyrészt az előbb említett pigment folt alatt és felett levő szaruhártya vastagsága eltérő: a felső (levegőn levő) szaruhártya vastagabb, így jobban véd mind az esetleges kiszáradástól, mind az UV sugárzástól. Másrészt (és ez az érdekesebb) a szemlencse nem gömb, hanem elnyúlt ovoid alakú: ez pedig azt teszi lehetővé, hogy mind a vízből, mind a levegőből érkező fénysugarak optimális görbületű lencsében törnek meg és alkotnak kvázi ideális képet a retinán.
(A képek az eBioMEDIA oldaláról származnak.)
Swamynathan SK, Crawford MA, Robison WG Jr, Kanungo J, Piatigorsky J. (2003) Adaptive differences in the structure and macromolecular compositions of the air and water corneas of the „four-eyed” fish (Anableps anableps). FASEB J. 17: 1996-2005.
 Egyéb közös vonások felemlegetése mellett, a gerincesek
Egyéb közös vonások felemlegetése mellett, a gerincesek  Mint azt mind a
Mint azt mind a  A kelet afrikai Nagytavakban (Malawi, Viktória és Tanganyika) élő sügérfajok mindig is az evolúció kirakatállatainak számítottak. A tavakban ma megfigyelhető több száz bölcsőszájú sügérfaj (cichlid) egyetlen közös ősből alakult ki, de mára a tavak biztosította összes élőhelyet kihasználják. Algaevő aljzatlakók és plaktonevő felszíni fajok színes kavalkádja jött létre élővilág egyik legnagyobb adaptív radiációja során.
A kelet afrikai Nagytavakban (Malawi, Viktória és Tanganyika) élő sügérfajok mindig is az evolúció kirakatállatainak számítottak. A tavakban ma megfigyelhető több száz bölcsőszájú sügérfaj (cichlid) egyetlen közös ősből alakult ki, de mára a tavak biztosította összes élőhelyet kihasználják. Algaevő aljzatlakók és plaktonevő felszíni fajok színes kavalkádja jött létre élővilág egyik legnagyobb adaptív radiációja során.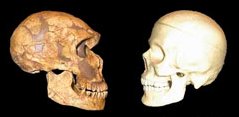 Az emberiség bölcsője Afrika, méghozzá (a jelenleg legelfogadottabb elmélet szerint) többszörösen is. A kétlábra emelkedő Homo genus először úgy 1 millió éve kolonizálta az eurázsiai kontinenst a Homo erectus formájában, majd ezt újabb hullámok követték. Először fél millió éve a Homo heidelbergensis egy csoportja kelt útra, majd úgy 75-100,000 éve a modern ember őse, a Homo sapiens.
Az emberiség bölcsője Afrika, méghozzá (a jelenleg legelfogadottabb elmélet szerint) többszörösen is. A kétlábra emelkedő Homo genus először úgy 1 millió éve kolonizálta az eurázsiai kontinenst a Homo erectus formájában, majd ezt újabb hullámok követték. Először fél millió éve a Homo heidelbergensis egy csoportja kelt útra, majd úgy 75-100,000 éve a modern ember őse, a Homo sapiens. A szépségnek ára van, még akkor is, ha bogarakról van szó. Pár hete már
A szépségnek ára van, még akkor is, ha bogarakról van szó. Pár hete már 
 Egy-egy bálnacsontvázzal szembesülve az átlagember első (és gyakran egyetlen) gondolata, hogy „hát igen, ez valóban bazi nagy”. Azonban a fizikai nagyság el- és felismerése mellett gyakran hajlamosak vagyunk elsuhanni az apróbb részletek mellett (persze néha kontár preparátorok is segítenek ebben).
Egy-egy bálnacsontvázzal szembesülve az átlagember első (és gyakran egyetlen) gondolata, hogy „hát igen, ez valóban bazi nagy”. Azonban a fizikai nagyság el- és felismerése mellett gyakran hajlamosak vagyunk elsuhanni az apróbb részletek mellett (persze néha kontár preparátorok is segítenek ebben). Állati szarvakról beszélve (már csak nyelvi okok miatt is) elsősorban mindenkinek a szarvas szokott beugrani, pedig az igazi nagymesterei a szarvnövesztésnek a bogarak között találhatók. A
Állati szarvakról beszélve (már csak nyelvi okok miatt is) elsősorban mindenkinek a szarvas szokott beugrani, pedig az igazi nagymesterei a szarvnövesztésnek a bogarak között találhatók. A 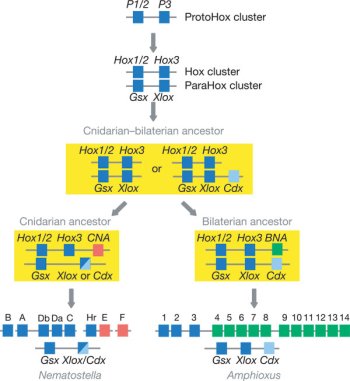 Ugyanis a csalánozó genomok segítségével rekonstruálhatjuk a Bilateria-Cnidaria csoportok szétválása előtti helyzetet (egyébként lásd
Ugyanis a csalánozó genomok segítségével rekonstruálhatjuk a Bilateria-Cnidaria csoportok szétválása előtti helyzetet (egyébként lásd