
 Mint arról bárki, aki vaksötétben próbált megtalálni egy kulcslyukat tanúskodhat, az éjszakai látás trükkös dolog. A hangsúly egyáltalán nem színeken van, hanem árnylatokon; valami észrevétele és meg nem látása közt alig néhány fotonnyi különbség van.
Mint arról bárki, aki vaksötétben próbált megtalálni egy kulcslyukat tanúskodhat, az éjszakai látás trükkös dolog. A hangsúly egyáltalán nem színeken van, hanem árnylatokon; valami észrevétele és meg nem látása közt alig néhány fotonnyi különbség van.
Érthető ennek függvényében, hogy az éjszakai életmódra berendezkedő állatok igyekszenek maximalizálni a retinájuk által felfogott fényt. Az ilyen irányú adaptáció egyik legismertebb következménye egy különleges fényvisszaverő réteg kialakulása a szemben – ez az ún. tapetum, és ez az oka például annak, hogy a macskafélék szeme sötétben világít.
Természetesen még létezhetnek finomítások, és egy ilyent a hetekben írtak le az Azara-éjimajom (Aotus azarae) (owl monkey) kapcsán.
Az Aotus génuszba tartozó éjimajmok egyik jellegzetessége, hogy az össze közeli rokonuk nappali, vagyis szinte biztos, hogy itt egy viszonylag a közelmúltban kialakult tulajdonsággal állunk szemben. Így elvileg az éjszakai látás élettani hogysmintjének megismerésén túl arra is fényt deríthetünk, hogy az evolúció milyen fejlődési folyamatokba „piszkált bele”.
Egy kapucinus majommal összehasonlítva, két dolog szinte dobja magát: egyrészt az Azara-éjimajom rendelkezik a fent említett tapetummal, másrészt jól látható, hogy a szemek mérete jelentősen megnőtt a koponyhoz képest, ami természetesen nem haszontalan, ha a vizuális információt igyekszünk maximalizálni. (Hogy részben lelőjem a poént: ez az a két tulajdonság amiről nem lesz szó a későbbiekben.) Van azonban egy harmadik, nagyon is lényeges változás, amihez némi szövettani viszgálódásra van szükségünk: az éjimajmok szemében a színérzékeny csapsejtek aránya, a monokromatikus, de fényre nagyon szenzitív pálcikákhoz képest lecsökkent (pontosabban arányiban több pálcikát lelünk).
A kérdés így arra egyszerűsíthető le (és itt tényleg átugrok néhány lépést), hogy milyen fejlődéstani lapja lehet egy ilyen változásnak? Miért és miként dönt úgy több sejt a retina kialakulása során, hogy pálcika-identitást választ?
A válaszhoz röviden meg kell ismerkednünk a retina kialakulásának egyik fejlődéstani szabályszerűségével: azok a sejtek amelyek a fejlődés korai szakaszaiban lépnek ki a sejtciklusból, azok csapok (cones) lesznek, míg a később kilépők pálcikák (rods). (Természetesen még számos más sejttípus van – kicsit bővebben lásd itt -, de a történet szempontjából most egyszerűsítenék.) És ez egyben sugal egy elég egyszerű evolúciós utat is az éjimajmok retinája felé: ha valami módon – pl. elhúzódik a sejtciklus – kevesebb sejt differenciálódik az egyedfejlődés korai fázisaiban, több sejt léphet ki a sejtciklusból később és válhat pálcika-fotoreceptorrá.
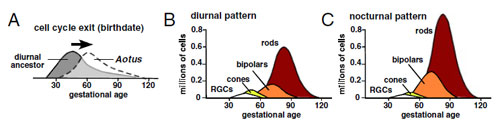 A jelek szerint az Aotus éjimajmok épp ezt a megoldást választották. (Persze ezt kimutatni egy kicsit trükkös, hiszen vizsgált, viszonylag közeli rokon majmok fejlődése is különbözik annyira, hogy nem hasonlítható össze csak a megtermékenyítéstől eltelt idő alapján.) A retina differenciácója az éjszakai fajokban (nocturnal) későbbre tolódott, és ez pont a számukra igen hasznos fényérzékeny sejttípus mennyiségét növelte meg.
A jelek szerint az Aotus éjimajmok épp ezt a megoldást választották. (Persze ezt kimutatni egy kicsit trükkös, hiszen vizsgált, viszonylag közeli rokon majmok fejlődése is különbözik annyira, hogy nem hasonlítható össze csak a megtermékenyítéstől eltelt idő alapján.) A retina differenciácója az éjszakai fajokban (nocturnal) későbbre tolódott, és ez pont a számukra igen hasznos fényérzékeny sejttípus mennyiségét növelte meg.
Dyer, M., Martins, R., da Silva Filho, M., Muniz, J., Silveira, L., Cepko, C., Finlay, B. (2009) Developmental sources of conservation and variation in the evolution of the primate eye. PNAS 106(22): 8963-8968. doi: 10.1073/pnas.0901484106
 A „
A „ Vannak olyan pillanatok, amikor egy addig számára ismeretlen fajt látva egy biológus szeme is nagyra tágul, és a hoaxra meg photoshopra gyanakodva kezd keresgélni az eredeti cikk után. Kb. ez volt esetemben is, amikor először megláttam a Macropinna microstoma fényképét.
Vannak olyan pillanatok, amikor egy addig számára ismeretlen fajt látva egy biológus szeme is nagyra tágul, és a hoaxra meg photoshopra gyanakodva kezd keresgélni az eredeti cikk után. Kb. ez volt esetemben is, amikor először megláttam a Macropinna microstoma fényképét. Nem várna el sokat az egyszeri olvasó, ha arra számítana, hogy 150 évvel „A fajok eredetének” megjelenése után, az evolúció körül zajló minden vita ellenére, maga a faj fogalma azért már kőbe véshetően letisztult lenne. Nincs így, és a rendelkezésünkre álló
Nem várna el sokat az egyszeri olvasó, ha arra számítana, hogy 150 évvel „A fajok eredetének” megjelenése után, az evolúció körül zajló minden vita ellenére, maga a faj fogalma azért már kőbe véshetően letisztult lenne. Nincs így, és a rendelkezésünkre álló  Annak, aki már töltött több-kevesebb időt valamely történeti múzeum falai között, annak talán fölösleges ecsetelni egy klasszikus páncélzat előnyeit és hátrányait. Előny, hogy megvéd a külvilág „kellemetlen” hatásaitól, hátránya viszont, hogy súlya és felülete függvényében, valamennyire mindenképpen korlátozza a mozgást.
Annak, aki már töltött több-kevesebb időt valamely történeti múzeum falai között, annak talán fölösleges ecsetelni egy klasszikus páncélzat előnyeit és hátrányait. Előny, hogy megvéd a külvilág „kellemetlen” hatásaitól, hátránya viszont, hogy súlya és felülete függvényében, valamennyire mindenképpen korlátozza a mozgást. Hogy gyors ütemben fussunk át a mindenki által egyébként is ismert alapokon: szimbiózisnak nevezzük az élővilág igazán hosszú távú kapcsolatait. Ezek egyik altípusa, a mindkét fél számára egyértelműen előnyös kapcsolat, az ún. mutualizmus. Ilyesmi figyelhető meg pl. egz korallzátonyon élő bohóchal és az őt körbevevő tengeri rózsák közt: míg utóbbiak csípős természetüknél fogva távol tartják a hal ellenségeit, addig a bohóchal elkergeti a tengeri rózsára kiéhezett halakat.
Hogy gyors ütemben fussunk át a mindenki által egyébként is ismert alapokon: szimbiózisnak nevezzük az élővilág igazán hosszú távú kapcsolatait. Ezek egyik altípusa, a mindkét fél számára egyértelműen előnyös kapcsolat, az ún. mutualizmus. Ilyesmi figyelhető meg pl. egz korallzátonyon élő bohóchal és az őt körbevevő tengeri rózsák közt: míg utóbbiak csípős természetüknél fogva távol tartják a hal ellenségeit, addig a bohóchal elkergeti a tengeri rózsára kiéhezett halakat. (Vendégmunkásunk, SexComb, újra lesújt.)
(Vendégmunkásunk, SexComb, újra lesújt.)
 Előre látott és kevésbé látott elfoglaltságok miatt, kicsit poszt-szegényebb időknek néz elébe a blog. De türelem, mert előbb vagy utóbb, visszatérünk a régi rendszereséghez. 🙂
Előre látott és kevésbé látott elfoglaltságok miatt, kicsit poszt-szegényebb időknek néz elébe a blog. De türelem, mert előbb vagy utóbb, visszatérünk a régi rendszereséghez. 🙂 Mint pár hete a Xiphophorus nemzettségbe tartozó halak, és a bennük levő onkogén kapcsán
Mint pár hete a Xiphophorus nemzettségbe tartozó halak, és a bennük levő onkogén kapcsán 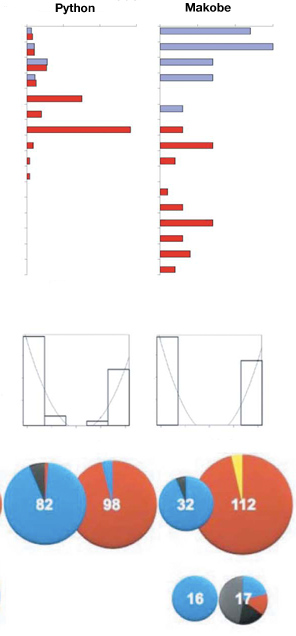 Hogy az elméletet teszteljék, közel ezer hímet vizsgáltak meg a Viktória-tó néhány szigete közelében. Az eredmény elég meggyőző. Először is, különösen a fokozatosan mélyülő helyeken, jól megfigyelhető, mint adja át a helyét a fémes-kék forma, a piros színűnek, a parttól egyre távolodva. Ha elég fokozatos a mélyülés (pl. Makobe estében – a függőleges tengely a mélységet jelöli), szinte nincs is átmeneti forma, csak a két „szélső érték” fedezhető fel, jól elkülönülve . (Ezt hivatottak ábrázolni a fekete-fehér oszlopdiagrammok a második sorban.)
Hogy az elméletet teszteljék, közel ezer hímet vizsgáltak meg a Viktória-tó néhány szigete közelében. Az eredmény elég meggyőző. Először is, különösen a fokozatosan mélyülő helyeken, jól megfigyelhető, mint adja át a helyét a fémes-kék forma, a piros színűnek, a parttól egyre távolodva. Ha elég fokozatos a mélyülés (pl. Makobe estében – a függőleges tengely a mélységet jelöli), szinte nincs is átmeneti forma, csak a két „szélső érték” fedezhető fel, jól elkülönülve . (Ezt hivatottak ábrázolni a fekete-fehér oszlopdiagrammok a második sorban.) Mióta magunk alakítjuk (egyre hatékonyabban) a körülöttünk levő világot, fokozatosan átalakult a „rátermettség” definíciója is, mind a saját, mind a hozzánk kapcsolódó élőlények viszonylatában.
Mióta magunk alakítjuk (egyre hatékonyabban) a körülöttünk levő világot, fokozatosan átalakult a „rátermettség” definíciója is, mind a saját, mind a hozzánk kapcsolódó élőlények viszonylatában.