
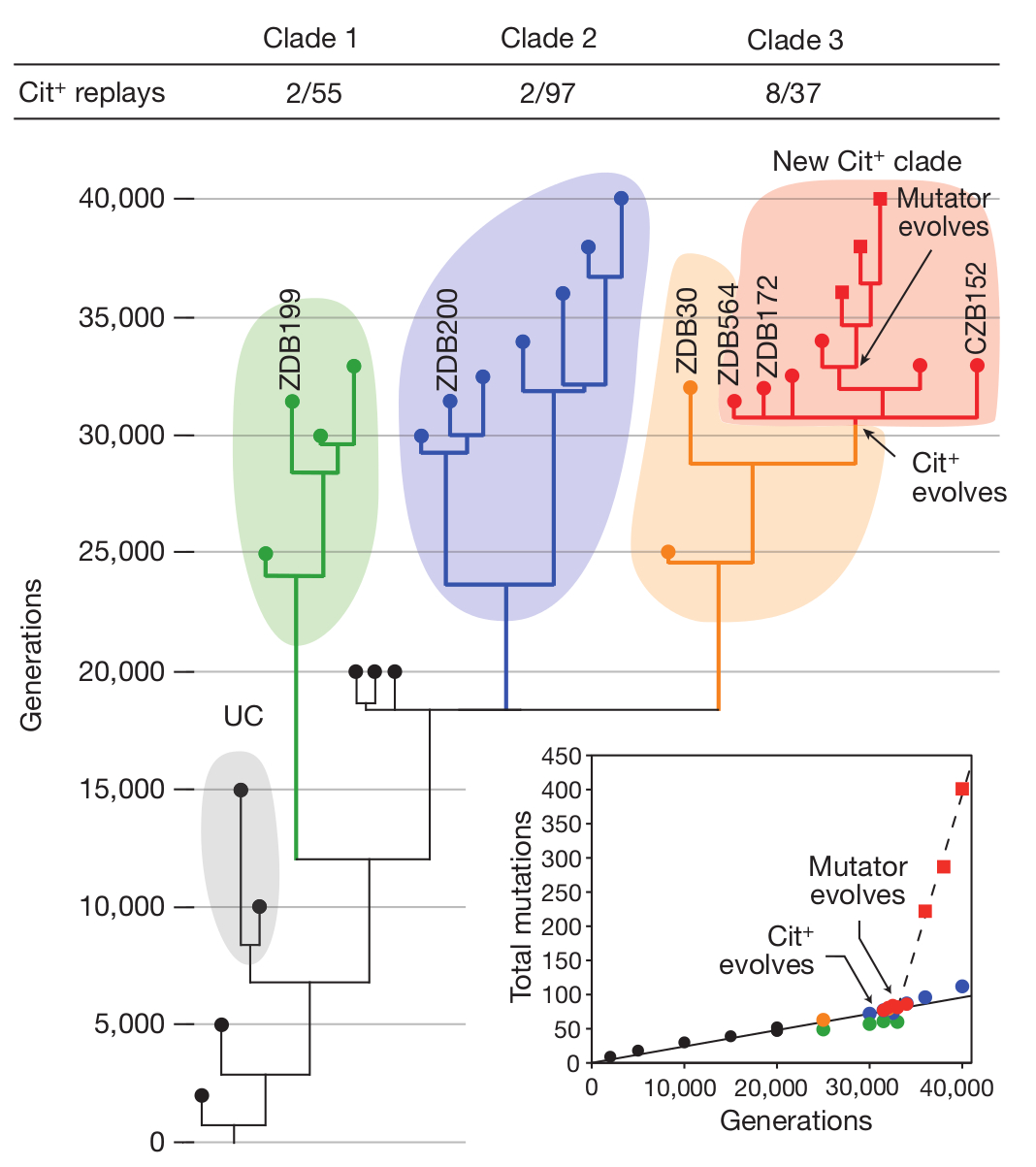
 Richard Lenskit nyilván senkinek sem kell bemutatnom, lebilincselően izgalmas kísérletei (1, 2, 3) elég sok adalékot szolgáltattak az evolúció megismeréséhez. Kétségkívül leghíresebb munkája az évtizedek óta folyó E. coli evolúciós kísérlete, ahol egy E. coli törzset többfelé osztott, majd szőlőcukorban szegény, citromsavban gazdag táptalajra helyezte őket és figyelte, mi történik velük. Az E. coli egyik jellemzője ugyanis, hogy csak anaerob körülmények között képes citromsavat hasznosítani, oxigén jelenlétében a citromsav nem jut át a sejtmembránon, így nem is képes táplálékként felhasználni a sejt. A mikrobiológiai gyakorlatban ez alapján azonosítják E. coliként. Lenski eredeti feltételezése az volt, hogy mivel az E. coli anaerob körülmények között képe citromsavat is hasznosítani, megfelelő körülmények között előbb-utóbb az evolúció során kialakul az oxigén jelenlétében való citromsavfogyasztás képessége is. Nem csalódott, röpke 31000 nemzedék után a tizenkét törzs egyikében meg is jelentek citromsavat is hasznosító sejtek, ezeket nevezte el Cit+ -nak.
Richard Lenskit nyilván senkinek sem kell bemutatnom, lebilincselően izgalmas kísérletei (1, 2, 3) elég sok adalékot szolgáltattak az evolúció megismeréséhez. Kétségkívül leghíresebb munkája az évtizedek óta folyó E. coli evolúciós kísérlete, ahol egy E. coli törzset többfelé osztott, majd szőlőcukorban szegény, citromsavban gazdag táptalajra helyezte őket és figyelte, mi történik velük. Az E. coli egyik jellemzője ugyanis, hogy csak anaerob körülmények között képes citromsavat hasznosítani, oxigén jelenlétében a citromsav nem jut át a sejtmembránon, így nem is képes táplálékként felhasználni a sejt. A mikrobiológiai gyakorlatban ez alapján azonosítják E. coliként. Lenski eredeti feltételezése az volt, hogy mivel az E. coli anaerob körülmények között képe citromsavat is hasznosítani, megfelelő körülmények között előbb-utóbb az evolúció során kialakul az oxigén jelenlétében való citromsavfogyasztás képessége is. Nem csalódott, röpke 31000 nemzedék után a tizenkét törzs egyikében meg is jelentek citromsavat is hasznosító sejtek, ezeket nevezte el Cit+ -nak.
 Repülni drága mulatság, nekünk embereknek is, de mindazon szerencsés vagy kevésbé szerencsés fajoknak is, akik erre a mutatványra mindenféle motorikus segédeszköz nélkül képesek.
Repülni drága mulatság, nekünk embereknek is, de mindazon szerencsés vagy kevésbé szerencsés fajoknak is, akik erre a mutatványra mindenféle motorikus segédeszköz nélkül képesek.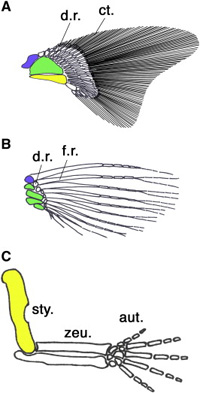 Amikor egyszer régen írtam egy
Amikor egyszer régen írtam egy 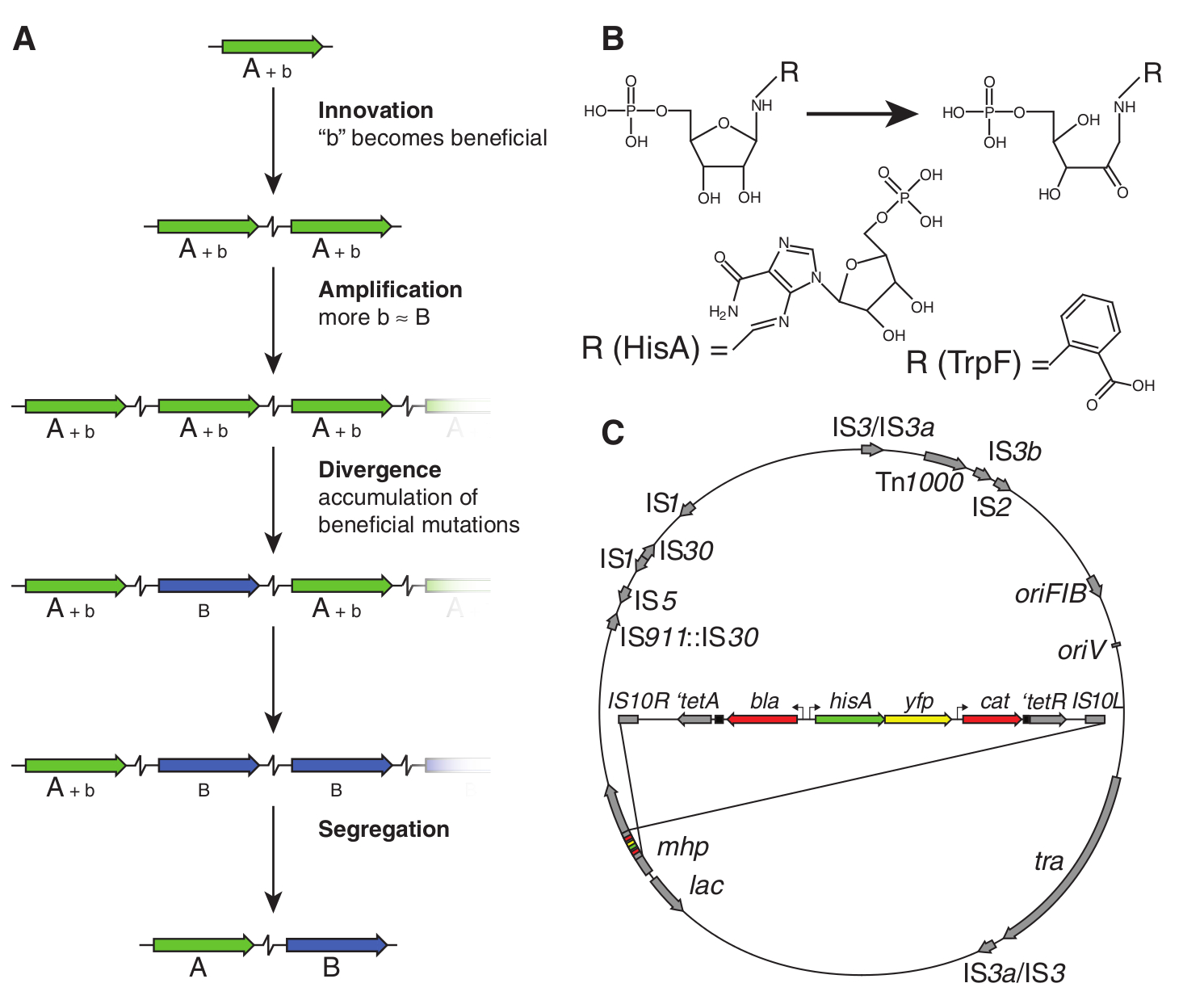
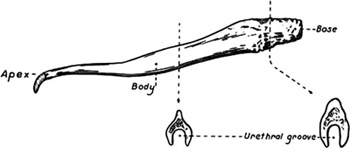 Kedvelt családi sztori szerint, néhány generációval ezelőtt, egyik női felmenő anatómia vizsgáján szóba került a férfi nemiszerv felépítése és a szigorú (naná, hogy férfi) vizsgáztató előtt fel kellett sorolni, hogy milyen szövetek hozzák létre a kényes jószágot. A végtelen zavarban levő vizsgázó el is kezdte sorolni: „bőr, izom, csont …” – „Azt maga csak úgy érzi!” csattant a közbevágás, és hát ez az a foka a megsemmisülésnek, ahonnan már nehéz visszahozni a dolgot.
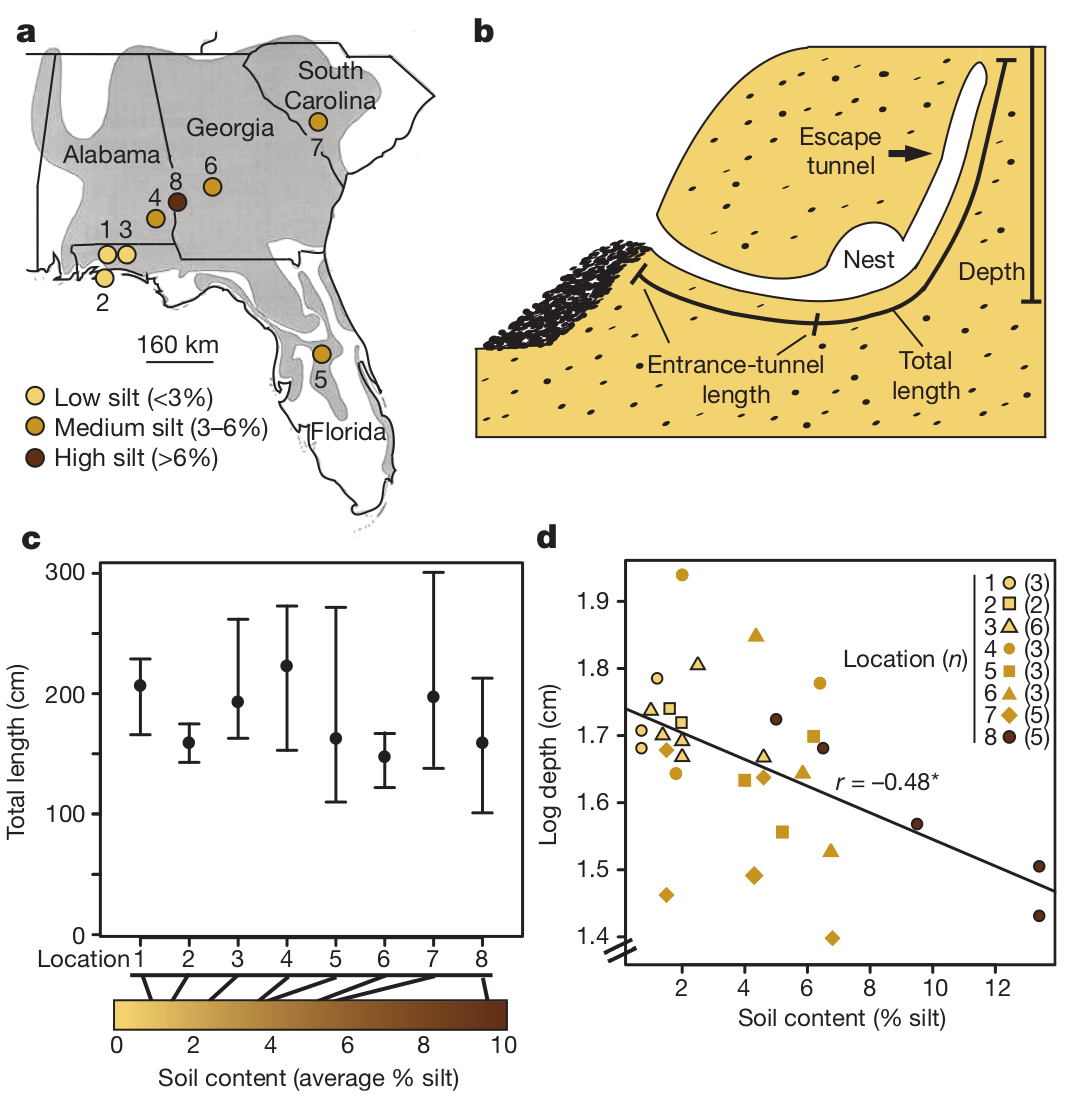
Kedvelt családi sztori szerint, néhány generációval ezelőtt, egyik női felmenő anatómia vizsgáján szóba került a férfi nemiszerv felépítése és a szigorú (naná, hogy férfi) vizsgáztató előtt fel kellett sorolni, hogy milyen szövetek hozzák létre a kényes jószágot. A végtelen zavarban levő vizsgázó el is kezdte sorolni: „bőr, izom, csont …” – „Azt maga csak úgy érzi!” csattant a közbevágás, és hát ez az a foka a megsemmisülésnek, ahonnan már nehéz visszahozni a dolgot. Kevés faj rendelkezik olyan formagazdagságal, mint a kutya (pontosabban a szürke farkas, hiszen ő a faj vadon élő ősi képviselője), az emlősök közül pedig jó eséllyel senki.
Kevés faj rendelkezik olyan formagazdagságal, mint a kutya (pontosabban a szürke farkas, hiszen ő a faj vadon élő ősi képviselője), az emlősök közül pedig jó eséllyel senki. A méhek kapcsán az utóbbi időben kicsit a
A méhek kapcsán az utóbbi időben kicsit a  És ha már konvergens evolúció, akkor itt egy másik remekbe szabott példa is – pontosabban csak részben új, hiszen, valami nagyon hasonlóról
És ha már konvergens evolúció, akkor itt egy másik remekbe szabott példa is – pontosabban csak részben új, hiszen, valami nagyon hasonlóról  Az elevenszülést általában az emlősökre jellemző tulajdonságnak tartjuk, pedig a legkülönbözőbb élőlénycsoportokban alkult ki egymástól függetlenül. Ma egy kevésbé ismert példáról lesz szó, a
Az elevenszülést általában az emlősökre jellemző tulajdonságnak tartjuk, pedig a legkülönbözőbb élőlénycsoportokban alkult ki egymástól függetlenül. Ma egy kevésbé ismert példáról lesz szó, a