
 Az elevenszülést általában az emlősökre jellemző tulajdonságnak tartjuk, pedig a legkülönbözőbb élőlénycsoportokban alkult ki egymástól függetlenül. Ma egy kevésbé ismert példáról lesz szó, a cecelégyről. A „cecelégy” alatt nem egyetlen fajt értünk, a Glossina genus körülbelül két tucat fajt tartalmaz, jelentősége nem is ebben rejlik, hanem hogy ezek a jószágok terjesztik az álomkórt, és egy rakás hozzá hasonló állatbetegséget, amelyek igen komoly problémát jelentenek a Szaharától délre. Mivel ilyen fontos kórokozók köztes gazdája, így folyamatosan keresik a módját, hogyan lehetne kiirtani, ezért viszonylag alaposan vizsgálták, így viszonylag pontosan ismerjük a szaporodását is.
Az elevenszülést általában az emlősökre jellemző tulajdonságnak tartjuk, pedig a legkülönbözőbb élőlénycsoportokban alkult ki egymástól függetlenül. Ma egy kevésbé ismert példáról lesz szó, a cecelégyről. A „cecelégy” alatt nem egyetlen fajt értünk, a Glossina genus körülbelül két tucat fajt tartalmaz, jelentősége nem is ebben rejlik, hanem hogy ezek a jószágok terjesztik az álomkórt, és egy rakás hozzá hasonló állatbetegséget, amelyek igen komoly problémát jelentenek a Szaharától délre. Mivel ilyen fontos kórokozók köztes gazdája, így folyamatosan keresik a módját, hogyan lehetne kiirtani, ezért viszonylag alaposan vizsgálták, így viszonylag pontosan ismerjük a szaporodását is.
Annak idején az iskolában azt tanultuk, hogy a rovarok petéket raknak, amikből lárvák kelnek ki. A cecelégy is valahogyan így kezdi, azonban egyszerre csak egyetlen egy pete érik meg a testében, ami a nőstény testén belül marad, a méhben, ahol a kikelő lárva az egész lárvális életét tölti. Amikor befejezte a lárvális fejlődést, az anyaállat kitojja a lárvát, ami azonnal bebábozódik, majd a bábból később kikel a kifejlett légy. Ebben az életciklusban a legszokatlanabb az az egészen nyilvánvaló tény, hogy a lárvának nincs ideje táplálkozni, az összes tápanyagot, ami egy kifejlett légy kikeléséhez szükséges az anyaméhen belül kell magához vennie. No de hogyan csinálja ezt a légy? A méh egy izmos falú, tracheákkal gazdagon behálózott falú szerv, ami elég nagy ahhoz, hogy egy harmadik lárvastádiumban lévő lárva is elférjen benne, amelynek a tömege megegyezik az anyaállatéval, hiszen a méhen kívül már nem táplálkozik, odabent kell elérnie a kifejlett testtömegét. A méhen belüli fejlődés meglepően hosszú, (A második ábrán, a nőstény ivarszerve látható az egyes időpontokban.) ez alatt az anyaállat a lárvát egy tejmiriggyel táplálja, amely a zsírtest közé benyomuló csőszerű mirigyekből áll. A mirigyek üregét kiválasztósejtek veszik körül, amelyek a tápláló tejet állítják elő a terhesség alatt. A cecelégy három különböző endoszimbionta baktériumot is hordoz, a Wolbachia pipientis (1, 2, 3) a petefészekben él és a petékkel együtt adódik át nemzedékről nemzedékre. A Sodalis glossinidius és a Wigglesworthia glossinidia is feltételezhetően a tejmirigyek váladékával kerül át az utódba. A tejmirigyek váladéka főleg fehérjéket és lipideket tartalmaz, két tejfehérje ismert, a tejmirigy fehérje (milk gland protein – GmmMGP) és a traszferrin (GmmTRF), más fehérjéket is kimutattak már, de ezek gyakorlatilag ismeretlenek. A szaporodás szempontjából fontos még a pete szikanyagát adó szikfehérje (yolk protein 1 – GmmYP1).
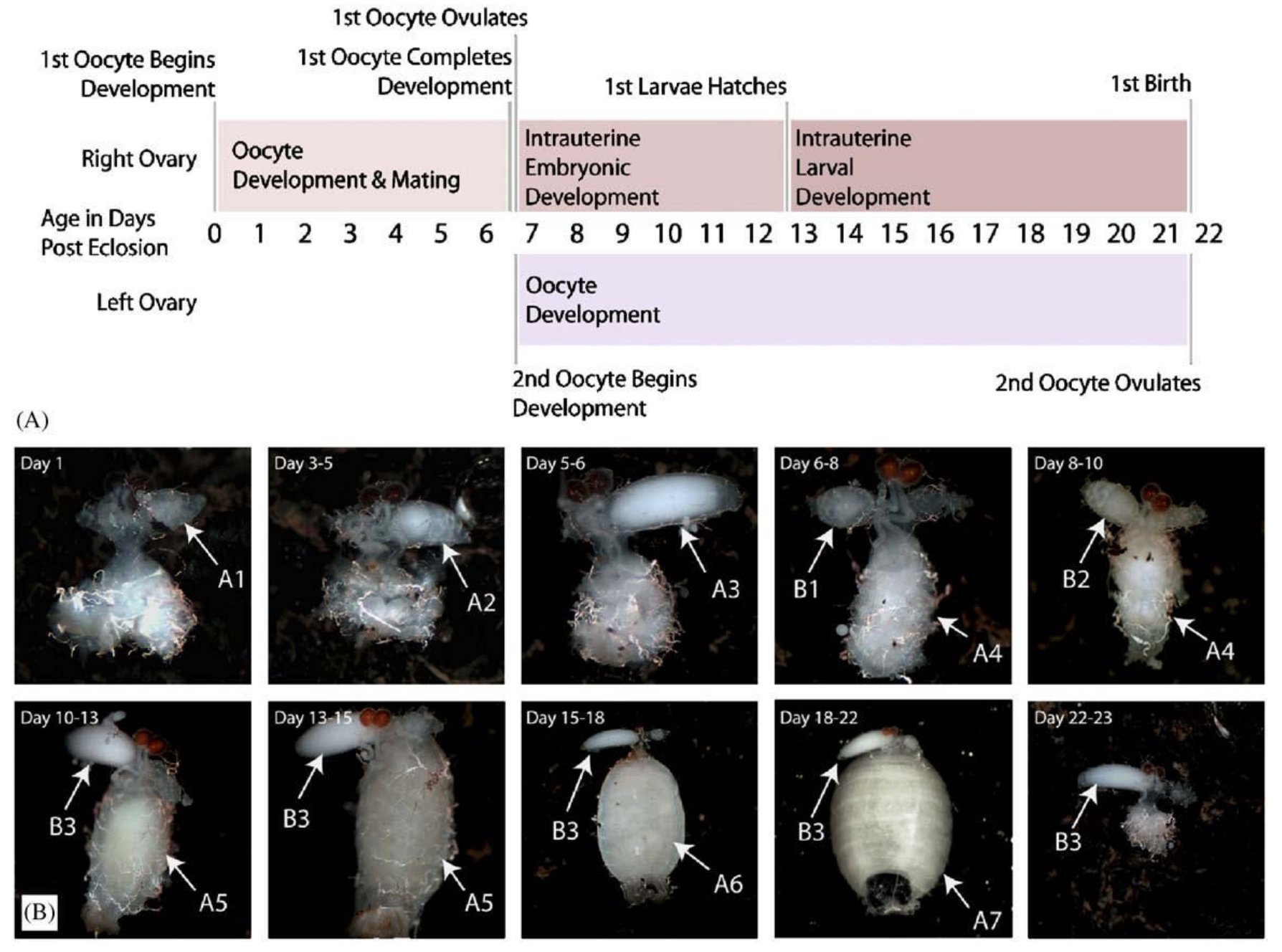
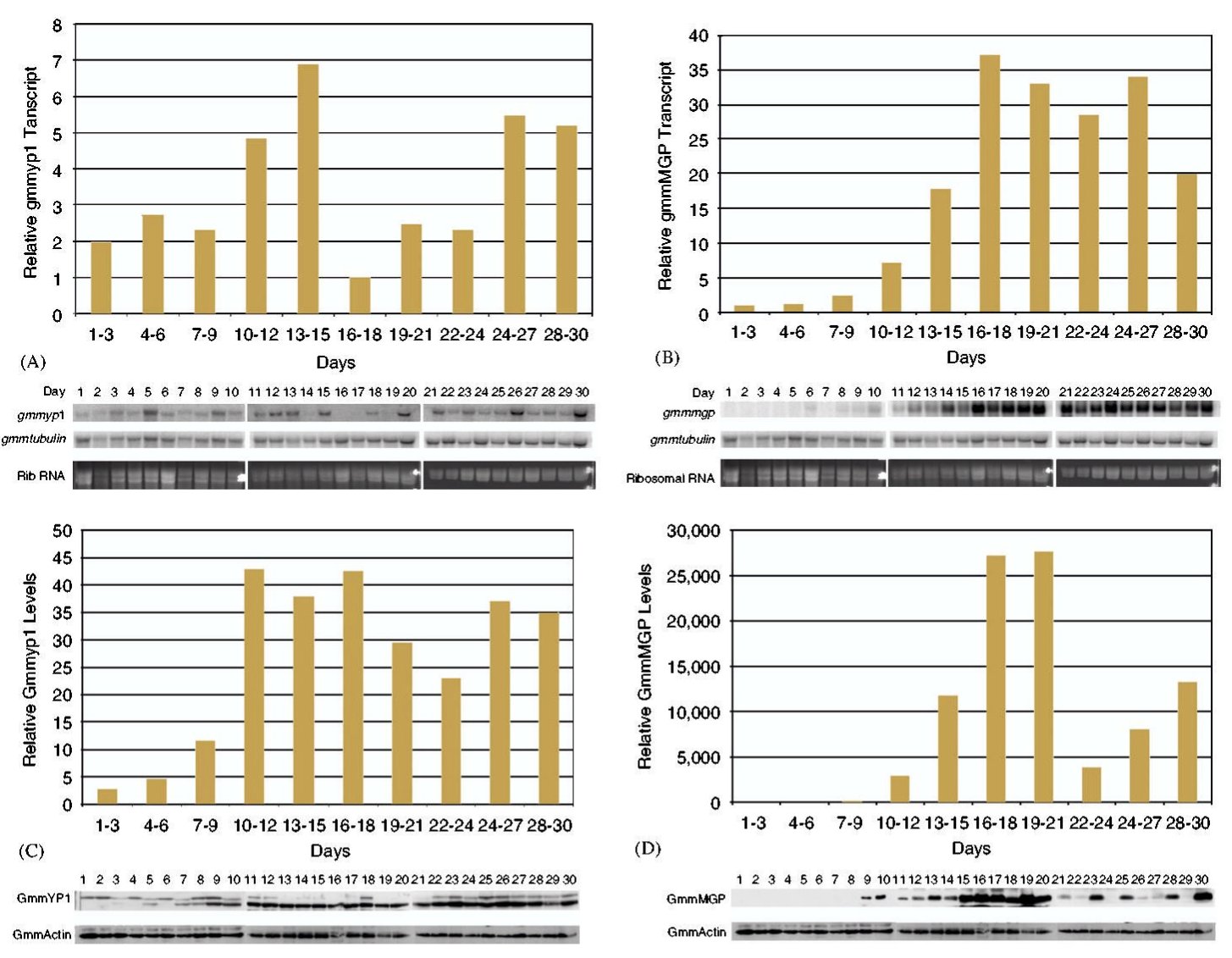
Mivel a szaporodásából következően a cecelégy nem túl szapora rovar, egy nőstény élete során 8-10 utódot képes csak kihordani, kézenfekvőnek tűnik, hogy az álomkór visszaszorításához vezető egyik fontos lépés lenne a cecelégy szaporodásának befolyásolása. Ez annál inkább lehetséges, mivel a nőstény cecelégy életében általában csak egyszer párzik, a spermiumokat eltárolja és azokkal termékenyíti meg a többi petét is. Geoffrey M. Attado és munkatársai azt vizsgálták, hogyan is működik a cecelégy szaporítószerve. Először northern blot módszerrel megvizsgálták, hogy a GmmMGP és a GmmYP1 gének kifejeződése hogy változik a szaporodási ciklus alatt, ez látható a harmadik ábrán (A és B), de érdekes módon western blot módszerrel azt is megvizsgálták, hogy mennyi fehérje transzlálódik a messenger RNSről (C és D). Nem meglepő módon a pete érése és az embrionális fejlődés során sok szikfehérje termelődik és kevés tejfehérje, míg a lárva kikelése után kevés szikfehérje és sok tejfehérje.
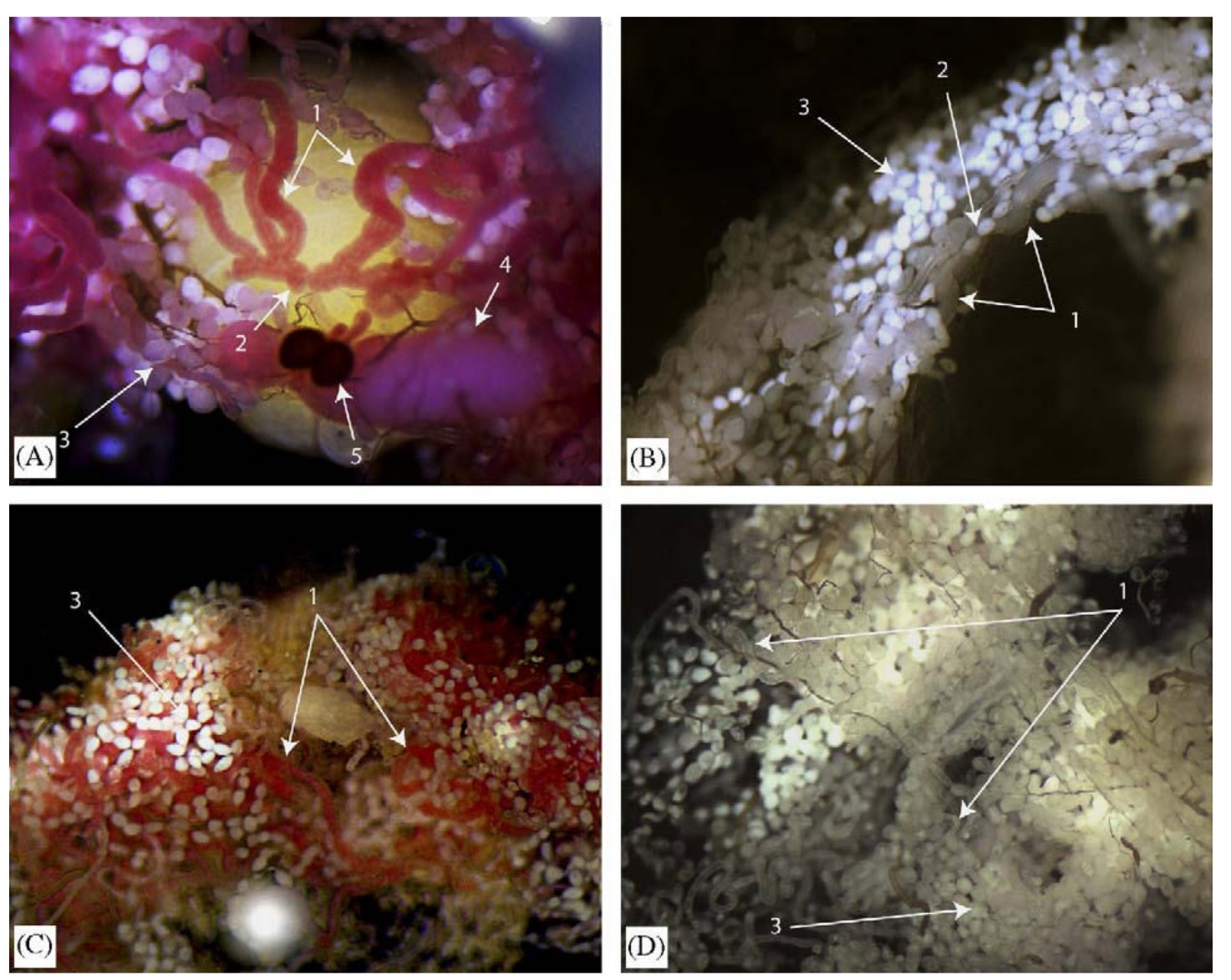
Nem meglepő módon azt tapasztalták, hogy az ivarszervekben fejeződik ki a szikfehérje, míg a tejmirigyekben a tejfehérje, ezt mutatja a következő ábra. itt az „1” nyíl mutat a tejmirigyekre, a „2” nyíl a méhre, a „3” nyíl a zsírtestre, a „4” nyíl a fejlődő petére, az „5” nyíl pedig a spermatékára. Az A és C ábra mutatja az immunhisztokémiás festést, a B és D kép a negatív kontroll, ahol immunizálatlan állat savóját használták elsődleges ellenanyagként.
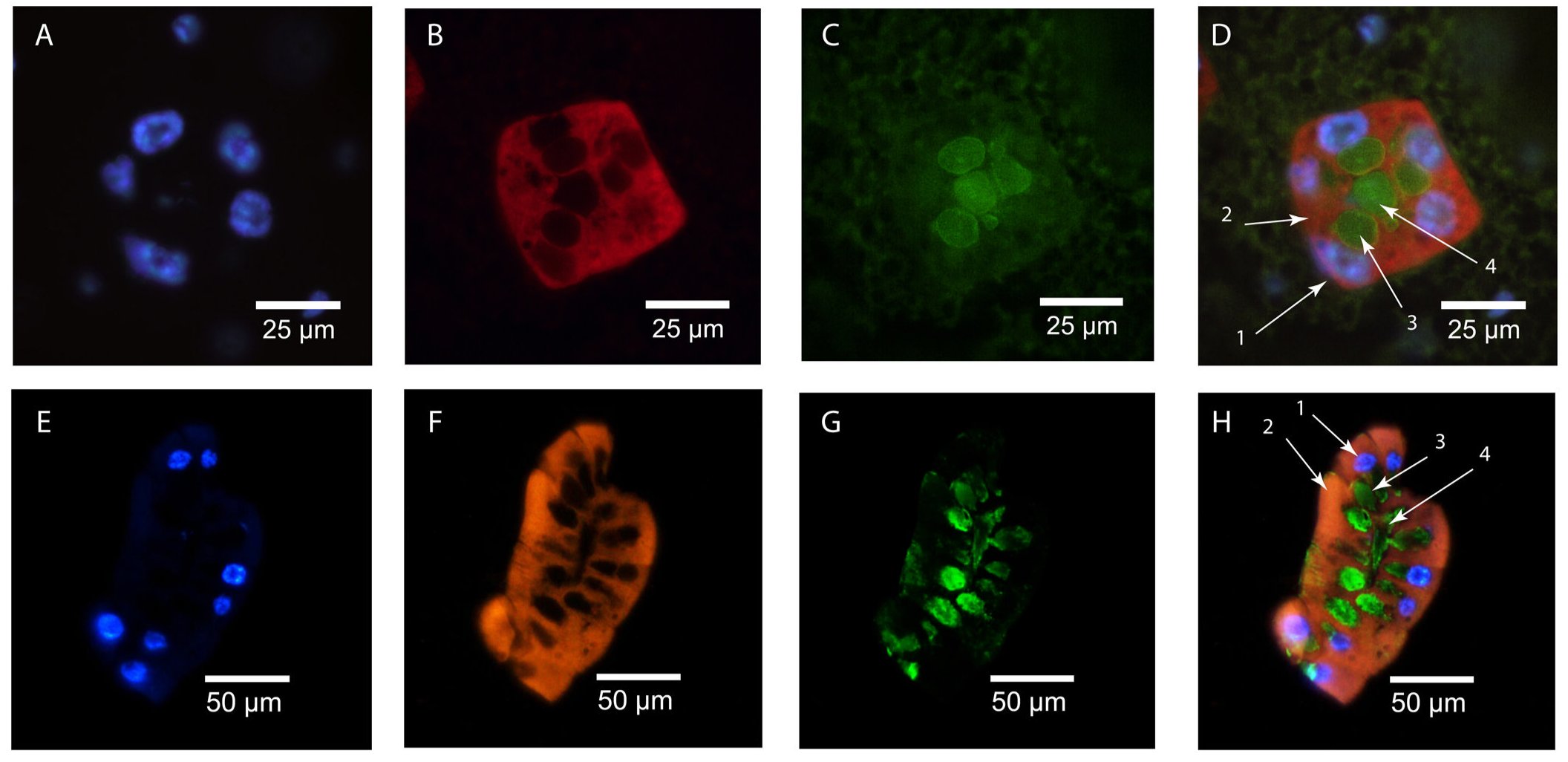
A következő cikkükben már kifinomultabb módszerekkel vágtak neki a vizsgálatoknak, pontosan meg tudták határozni, hogy fejeződik ki a gmmmgp gén és hol található a GmmMGP fehérje. Ezt mutatja a következő ábra. A fölső sor a keresztmetszeti kép, az alsó a hosszmetszeti. A kék egy DNS festék (DAPI), ami a sejtmagokat jelöli ki, a narancssárga a gmmmgp RNS -t jelöli ki in situ RNS hibridizálással, a zöld a GmmMGP fehérje immunfluoreszcens festésének az eredményét mutatja. Világosan látszik, hogy a tejfehérjét a tejmirigy üregét körülvevő kiválasztósejtek termelik, bennük fejeződik ki a gmmmgp gén, majd a fehérjét a tejmirigy üregébe továbbítják. Ugyanígy megvizsgálták a transzferrin fehérje kifejeződését is, hasonló festődést tapasztaltak, a transzferrint is a kiválasztósejtek termelik és a tejmirigy üregébe juttatják.
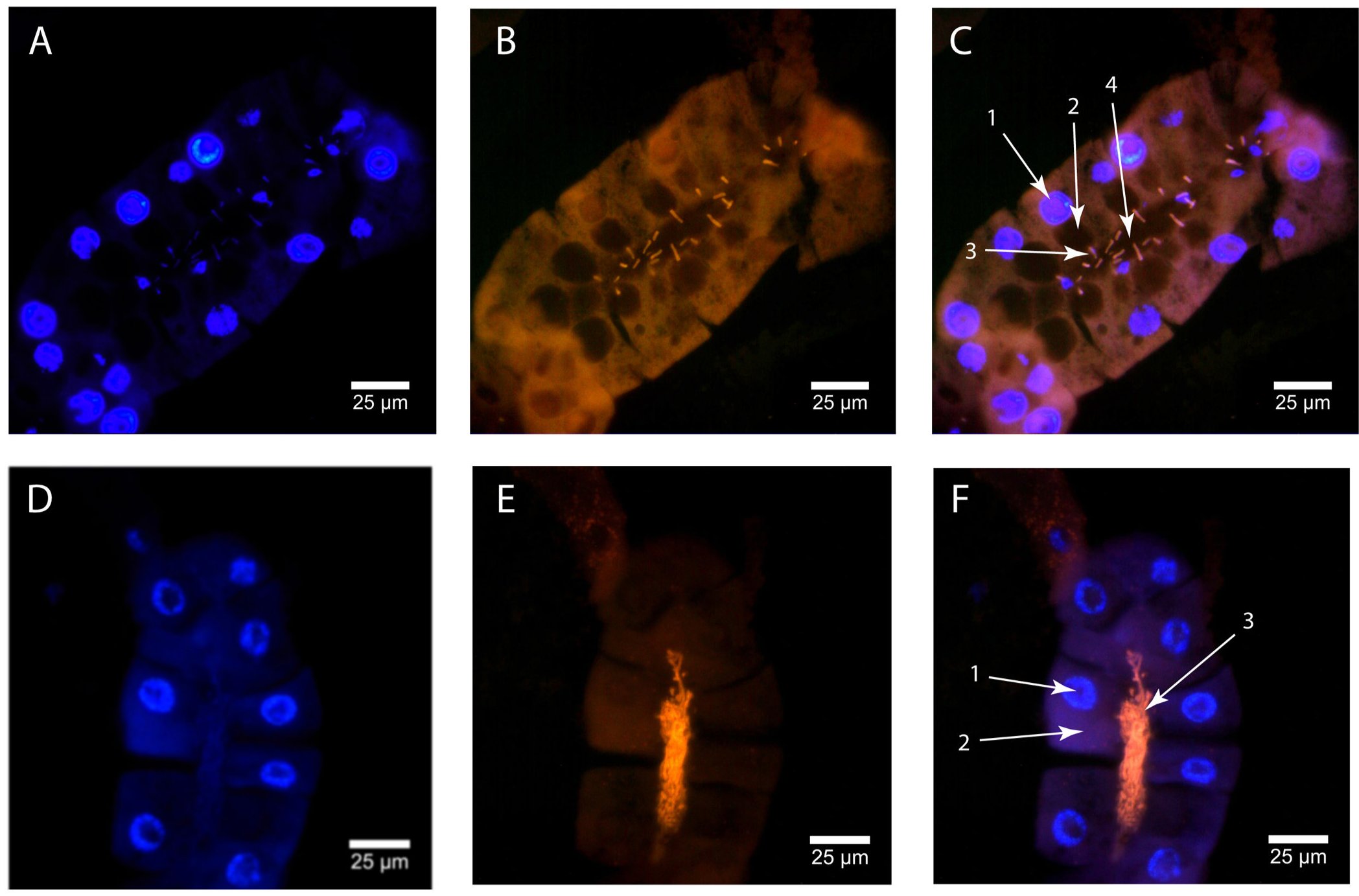
RNS inhibícióval lecsökkentették a gmmmgp RNS mennyiségét a nőstényekben, aminek hatására a vizsgált egyedek feleannyi utódot hoztak világra, mint a kontroll kettősszálú RNS -sel kezelt állatok (a két csoport által lerakott bábok számát mutatja a következő ábra), ebből arra következtettek, hogy a tejfehérje szükséges az utód kifejlődéséhez.
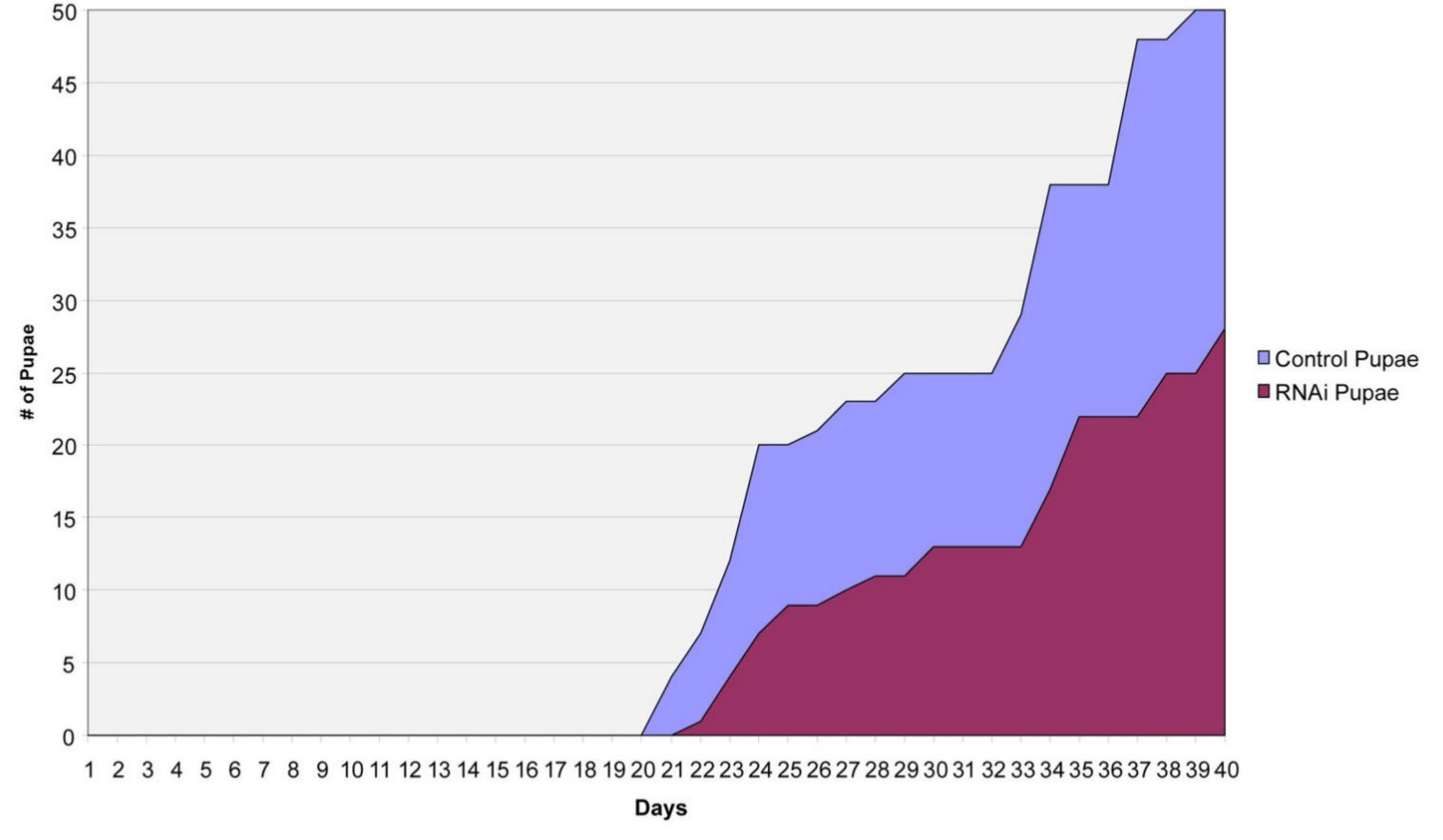
Megvizsgálták a szimbionta baktériumok eloszlását is a tejmirigyekben, ez látható a következő ábrán. Azt tapasztalták, hogy a Sodalis sejtek a tejmirigy üregében és a kiválasztósejtek citoplazmájában is kimutathatóak, míg a Wigglesworthiák kizárólag a tejmirigyek üregében láthatóak.
A Sodalis endoszimbionták eloszlása:
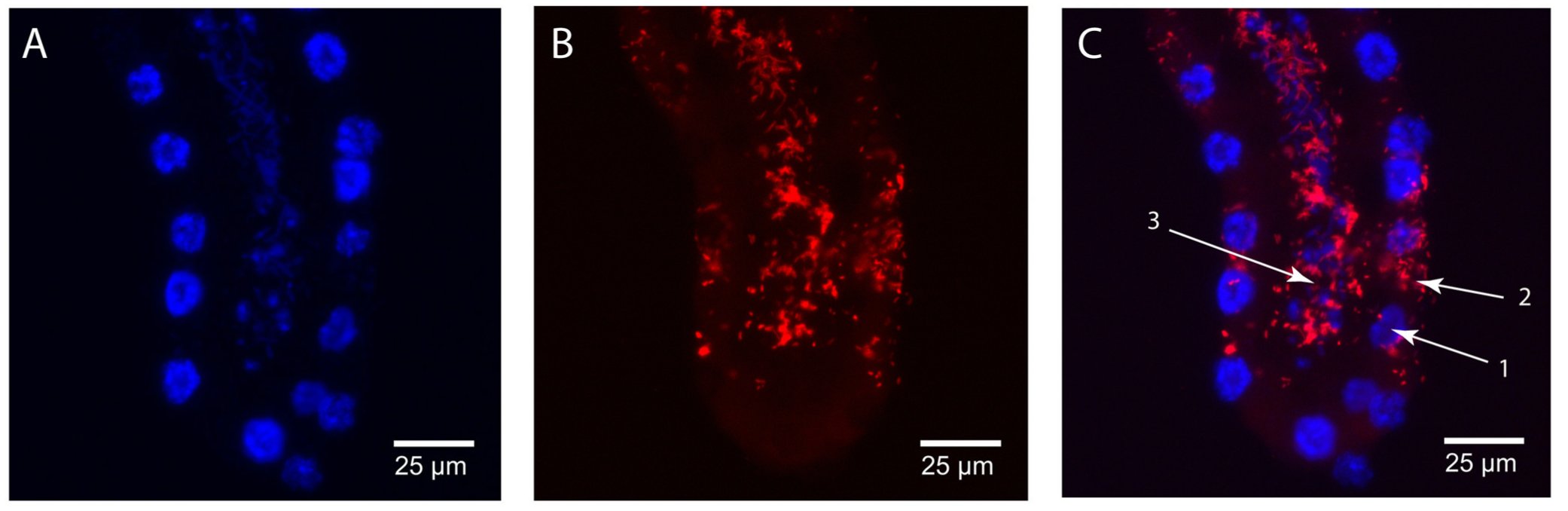
A Wiggleswothiák eloszlása:
A cecelégy szaporodása mindenképpen szokatlan az ízeltlábúak között, nyilván az teszi lehetővé ezt a roppant energiaigényes terhességet, hogy a légy nagy termetű emlősállatok vérével táplálkozik, így elég tápanyaghoz jut, hogy ellássa az utódait egész lárvális életükben. Egyelőre elég kevéssé ismerjük még ennek a szaporodási módnak a hátterét, de amint látható, vannak akik ezt a jószágot kutatják.
Attardo GM, Guz N, Strickler-Dinglasan P, Aksoy S. (2006): Molecular aspects of viviparous reproductive biology of the tsetse fly (Glossina morsitans morsitans): regulation of yolk and milk gland protein synthesis. J Insect Physiol. 2006 52(11-12):1128-36.
Attardo GM, Lohs C, Heddi A, Alam UH, Yildirim S, Aksoy S. (2008): Analysis of milk gland structure and function in Glossina morsitans: Milk protein production, symbiont populations and fecundity. J Insect Physiol. 54(8):1236-42
„…pedig a legkülönbözőbb élőlénycsoportokban fejlődött ki egymástól függetlenül.”
Ez a sor tudománytalan. A fejlődésre semmilyen bizonyíték nincs.
Az egymástól való függetlenségre sem.
„A cecelégy szaporodása mindenképpen szokatlan az ízeltlábúak között”
Talán a cecelégy (is) tervezett?
@ID_EGEN:
Kijavítottam „kialakulásra”.
Akkor ha valami szokatlan, az tervezett? Ami megszokott, az nem? Akkor az összes többi légy, olyan százhúszezer faj nem tervezett? Csak ez a pár cecelégy?
Mi az ami szokatlan, és mi a megszokott? A cecelégy szaporodása eltérő a többi tervezett ízeltlábúétól.
Ha valami létezik, van, arról azt is fel kell tételezni, hogy tervezett, vagy tervezve alakult ki, azt viszont hogy egymásból spontán alakultak ki, súlyos tudománytalanság.
Magyarul ami úgy szaporodik, mint a többi légy, az tervezett. Ami nem, az is. Akkor ennek mi köze a szaporodáshoz? Ez alapján nem lehet különbséget tenni a tervezett és a nem tervezett élőlények közt.
Re: ID_EGEN@1012. 07. 11. 20:48
Ha az az alapfeltevés minden létező valamire, hogy az tervezett, abból milyen közetkeztetést lehet levonni azon kívül hogy „létezik (legalább) egy ‘tervező'”? (Amiből mellesleg az is következik, hogy a tervező is tervezett — tovább folytatva a gondolatmenentet, feltéve, hogy valami nem tervezheti meg ön-magát, eljutunk tervezők végtelen sorához, amit valamiért nem szerettek.) Mit lehet ebből megtudni a tervezö(k)ről? Merthogy ez lenne az egész „tudományosdi”nak a célja: minél többet megtudni a világról amiben élünk: és lám, van egész jó orvostudományunk (többek között), ami a biológián alapul (az evolució elméletét is felhasználva).