
 És ha már konvergens evolúció, akkor itt egy másik remekbe szabott példa is – pontosabban csak részben új, hiszen, valami nagyon hasonlóról már írtunk egyszer.
És ha már konvergens evolúció, akkor itt egy másik remekbe szabott példa is – pontosabban csak részben új, hiszen, valami nagyon hasonlóról már írtunk egyszer.
Konkrétabban arról van szó, hogy két, egymással csak távoli rokonságban álló csoportban, a cetekben és denevérekben – amelyek egymástól függetlenül a tájékozódásnak hasonló formáját választották, az echolokációt – nagyon hasonló molekuláris változások következtek be a belső fül hangérzékelésért felelős komponenseiben.
Mivel esetleg már nem mindenki emlékezik megfelelő mélységben a gimnáziumi biológia tananyag hallást elemezgető részeire, kezdjünk egy gyors összefoglalással, miként is működik a hallás.
A környezetünkből érkező hangullámok először a dobhártyát rezegtetik meg, majd ez a rezgés a kis hallócsontocskák révén átadódik a belső fül folyadékkal töltött és csigaház alakú szervének, a nagyon eredeti módon csigának (latinul cochlea) nevezett struktúrának.
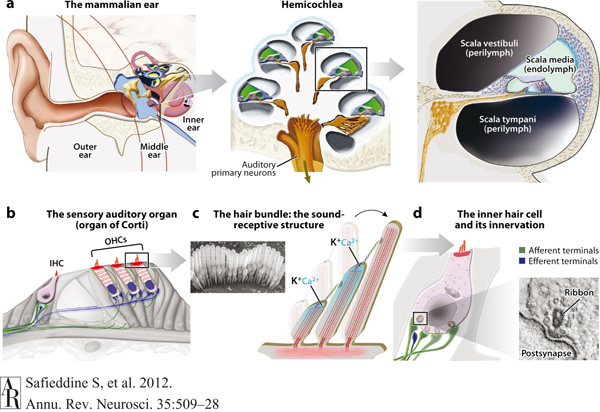
A csigában szendvics módon két perilimfatikus folyadékkal teli tér veszi körül a hallás valódi helyét, az ún. scala mediát (amely egy másik folyadékkal, az endolimfával van töltve). A perilimfával teli részből a rezgés átadódik az endolimfának, és itt az ún. Corti szervben a különleges hallósejtek alakítják át idegi ingerré a fizikai jelet.
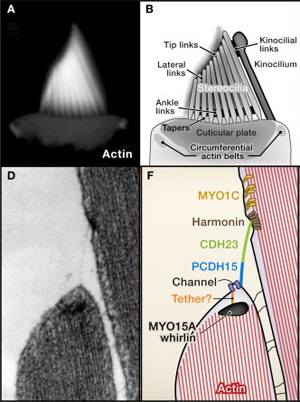 Maga az átalakítás is érdemel néhány szót: a szóban forgó szőrsejtek felszínén növekvő méretű kis kitüremkedések láthatóak, pánsípra emlékeztető elrendeződésű kinövések. Ezeknek a teteje apró molekuláris láncokkal van összekötve, amelynek köszönhetően, az egy-egy sejthez tartozó összes „szőr” egyszerre fog hajolni, amikor az endolimfa rezgésbe jön. A molekuláris láncok húzóereje azonban azt is okozza, hogy a szőrök felszínén levő apró ioncsatornák kinyílnak, s ezáltal a rezgés hatására megváltozik a sejtmembrán polarizációja. Jel keletkezik, ami aztán továbbítódik az agyunkba.
Maga az átalakítás is érdemel néhány szót: a szóban forgó szőrsejtek felszínén növekvő méretű kis kitüremkedések láthatóak, pánsípra emlékeztető elrendeződésű kinövések. Ezeknek a teteje apró molekuláris láncokkal van összekötve, amelynek köszönhetően, az egy-egy sejthez tartozó összes „szőr” egyszerre fog hajolni, amikor az endolimfa rezgésbe jön. A molekuláris láncok húzóereje azonban azt is okozza, hogy a szőrök felszínén levő apró ioncsatornák kinyílnak, s ezáltal a rezgés hatására megváltozik a sejtmembrán polarizációja. Jel keletkezik, ami aztán továbbítódik az agyunkba.
(Mivel nem tartozik szorosan a témához, nem megyek bele a részletekbe, de nagyon fontos magyar vonatkozása van, hogy miért halljuk különbözőnek az eltérő frekvenciájú hangokat – akit érdekelnek a részletek, olvassa a Nobel díjas Békésy György visszaemlékezését híres felfedezésére.)
A rendszer felépítése jól ismert tehát, miként működésének logikája is, de azt még csak tanulgatjuk, hogy milyen módon optimalizálható a rendszer a különböző extrém-hallástípusokra, pl. az echolokációra.
Márpedig a jelek szerint van ilyenfajta optimalizáció, ezt bizonyította a már korábban említett, a szőrsejtek membránjában levő Prestin nevű fehérje konvergens evolúciója cetekben és denevérekben, és ezt bizonyítja három újabb hallás-fehérje konvergens evolúciója a két csoport között.
A háromból kettő, a Cdh23 és a Pcdh15, az előbb bemutatott, egymás melletti „szőröket” összekötő molekulárs lánc két fontos eleme, a harmadik pedig, az Otof nevű fehérje, ami a szőrsejt különleges szinapszisában (azon a felületen, ahol a kapcsolódó idegsejtnek átadja az ingerületet) játszik szerepet.
Mindhárom esetben valami nagyon hasonló figyelhető meg (alább a Cdh23 esetében látható a dolog): ha a fehérjét kódoló gének nukleoitsorrendjének hasonlósága alapján szerkesztünk törzsfát, akkor az hűen fogja tükrözni az állatok „hivatalos” leszármazási viszonyait. A piros és kék színnel jelölt rokon, és egyaránt echolokációval tájékozódó csoportok viszonylag közel lesznek egymáshoz (a Hipposideros nemzettségbe tartozó fajok más óvilági, nem echolokáló gyümölcsevő denevérkkel mutatják a legközelebbi rokonságot), a delfin (Tursiops truncatus) pedig távolabb esik. Ha azonban a fehérjék aminosavsorrendje alapján szerkesztjük a törzsfát, teljesen más képet kapunk: az echolkáló fajok mintha közeli rokoságban állnának egymással.

Ezek alapján nem túlzás azt feltételezni, hogy a fehérjék szekvenciája az echolokációhoz szükséges optimum felé változott mindhárom csoportban, számos hallásban fontos fehérje esetében. A nagy kérdés és erre sajnos még nincs válasz, hogy miért optimáisabb ez a szekvencia az echolokációhoz.
Shen Y-Y, Liang L, Li G-S, Murphy RW, Zhang Y-P (2012) Parallel Evolution of Auditory Genes for Echolocation in Bats and Toothed Whales. PLoS Genet 8(6): e1002788. doi:10.1371/journal.pgen.1002788
Gillespie PG, Müller U (2009) Mechanotransduction by hair cells: models, molecules, and mechanisms. Cell 139(1): 33-44.
Safieddine S, El-Amraoui A, Petit C (2012) The auditory hair cell ribbon synapse: from assembly to function. Annu Rev Neurosci 35: 509-28.
Egy érdekes eset: vak srác csettintéssel tájékozódik.
http://www.youtube.com/watch?v=qLziFMF4DHA