
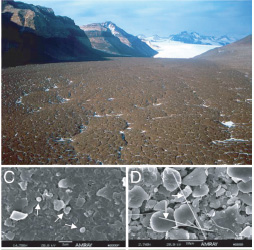
 Az egyik dolog amitől a napokban zengett a (tudományos) sajtó, az az antarktiszi jégmintákból kitenyésztett mikróbákhoz fűződik (a másikról kicsit később).
Az egyik dolog amitől a napokban zengett a (tudományos) sajtó, az az antarktiszi jégmintákból kitenyésztett mikróbákhoz fűződik (a másikról kicsit később).
Személyszerint picit vitatnám azokat az állításokat , hogy a százezer ill. nyolcmillió éves mintákból kinyerni élő anyagot totál meglepő lenne – hiszen hasonló kunsztot már véghezvittek 200 millió éves sókristályokkal is -, de persze az valahol természetes, hogy a szerzők igyekeznek nagy feneket keríteni saját cikküknek (de ettől még nem kellene minden szuperlatívuszt gondolkodás nélkül bevenni).
Viszont a fenti kekeckedésemtől függetlenül, vagy épp azzal együtt, az eredmény érdekes. A két említett mintában csak baktériumok vannak jelen (archebaktériumoknak és eukariótáknak nem lelték nyomát), és azok is viszonylag fajszegényen: a fiatalabb mintában leginkább Arthrobacterekkel rokon fajok domináltak, míg az idősebben Leptothrix fajok.
Az talán kevésbé volt meglepő, hogy a százezer éve "lefagyasztott" bacik jobb bőrben voltak (azért a DNS a javító mechanizmusok nélkül roncsolódik az idők során, még fagypont alatt is), s ennek megfelelően kb. 7 nap alatt tudták számukat megduplázni, szemben 30-70 nappal, amire nyolcmillió éves rokonaiknak volt szükségük. Az viszont inkább, hogy mindkét minta esetében az izolált DNS szakaszok ~40%-a nem hasonlított semmire a bakteriális genom-adatbázisokban. Ennek egyik magyarázata persze az is lehetne, hogy a DNS teljesen szétmutálódott/roncsolódott, de ez persze elég kézenfekvő volt ahhoz, hogy megfelelő ellenőrzásekkel kiküszöbölhető legyen. Vagyis úgy tűnik, hogy egy csomó új génnel állunk szemben. Pár éve ez még elképesztően izgalmasnak tűnt volna, de azóta jobban tudatában vagyunk saját ismereteink határainak mikrobiális diverzitás terén: a közelmúltban befejezett (kisléptékű) óceáni mintavételek 1700 új fehéjecsaládot írtak le! (Hiába no, többfajta bacik vannak földön és égen, Horatio…)
És mivel ez az régi-új genetikai sokféleség olvadó jégrétegekben fekszik, az is elképzelhető, hogy ha a gleccserek olvadásával ezek az ősi bacik bejutnak a tengerekbe és kapcsolatba kerülnek mai társaikkal, akkor a horizontális géntranszfer nevű mechanizmus révén átadhatják nekik ősi géneiket, vagyis azokat gyorsítópályán küldhetik vissza a természetes szelekció "tesztmezejére".
Mindenesetre, ha egy-egy baci több millió évig képes jégbefagyva kihúzni ennyire mostoha körülmények között, akkor persze azon is el lehet filózni, hogy ezt csak a Földön tudná, vagy pl. egy üstökös belsejében is… A pánspermia szimpatizánsai újabb érvvel gazdagodtak. 😉
Bidle KD, Lee SH, Marchant DR, Falkowski PG (2007) Fossil genes and microbes in the oldest ice on Earth. PNAS 104: 13455-13460.
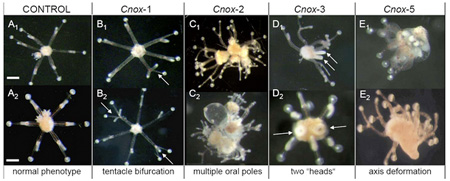 A Hox-gének,
A Hox-gének,  Nnna, akkor (egyébként nem létező) "Önök kérték" rovatunk keretében lássunk egy igazán érdekes problémát: a pókhálószövés rejtelmeit.
Nnna, akkor (egyébként nem létező) "Önök kérték" rovatunk keretében lássunk egy igazán érdekes problémát: a pókhálószövés rejtelmeit.

 Ha csak annyit írnék, hogy a posztban egy erősen szociális fajról lesz szó, melynek tagjai az egymással való érintkezéshez a kommunikáció egy igen fejlett formáját használják, és amely faj Afrikából (több hullámban) kiindulva hódította meg a világot, akkor valószínűleg sokan a Homo sapiens-re gondolnának,
Ha csak annyit írnék, hogy a posztban egy erősen szociális fajról lesz szó, melynek tagjai az egymással való érintkezéshez a kommunikáció egy igen fejlett formáját használják, és amely faj Afrikából (több hullámban) kiindulva hódította meg a világot, akkor valószínűleg sokan a Homo sapiens-re gondolnának,  Bő másfél évvel a
Bő másfél évvel a 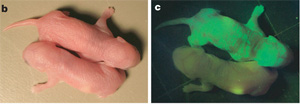 Mivel őssejtekről már amúgy is régen esett szó: szóval az a nagy hír, hogy végre-valahára úgy tűnik, hogy (legalább részben) sikerült az ún. transzdifferenciálódás kérdést megoldani. A beavatatlanok számára röviden összefoglalva ez az a jelenség, amikor egy véglegesen differenciálódott sejtből (pl. bőrsejt) – vagy annak már jócskán differenciálódott elődjéből – egy teljesen más fajta sejtet állítunk elő, olyant ami egy másik
Mivel őssejtekről már amúgy is régen esett szó: szóval az a nagy hír, hogy végre-valahára úgy tűnik, hogy (legalább részben) sikerült az ún. transzdifferenciálódás kérdést megoldani. A beavatatlanok számára röviden összefoglalva ez az a jelenség, amikor egy véglegesen differenciálódott sejtből (pl. bőrsejt) – vagy annak már jócskán differenciálódott elődjéből – egy teljesen más fajta sejtet állítunk elő, olyant ami egy másik 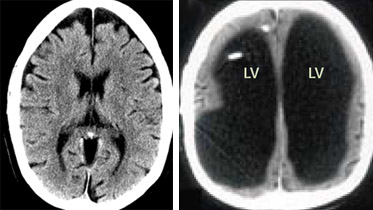
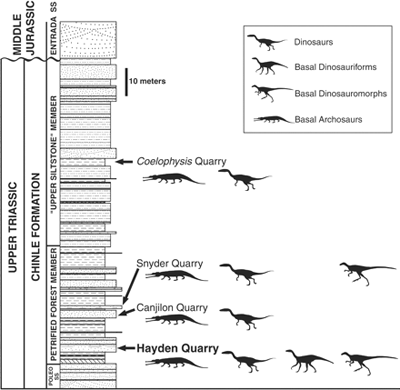 A dinoszauruszok a késői Triász folyamán jelentek meg (uszkve 200-230 millió éve) és ezt követően terjedtek el világszerte, fokozatosan kiszorítva más, primitívebb
A dinoszauruszok a késői Triász folyamán jelentek meg (uszkve 200-230 millió éve) és ezt követően terjedtek el világszerte, fokozatosan kiszorítva más, primitívebb 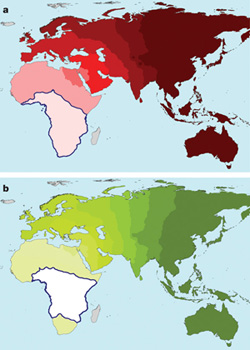 Akik járatosak a témában, biztos felfigyeltek rá, hogy mikor a múlt héten az emberi sokféleség eredetét (pontosabban amit erről ma tudunk/gondolunk)
Akik járatosak a témában, biztos felfigyeltek rá, hogy mikor a múlt héten az emberi sokféleség eredetét (pontosabban amit erről ma tudunk/gondolunk) 