

Már régóta foglalkoztatja a történészeket, hogy vajon létezett-e, létezhetett-e kapcsolat a dél-amerikai kontinens és a polinéz szigetvilág lakói között, a Kolumbusz előtti időkben. Számos elmélet és cáfolat született eddig ezügyben (a leghíresebb talán Thor Heyerdahl Kon-Tiki expedíciója), de igazán közvetlen bizonyíték eddig nem került elő. Márpedig DNS-bizonyíték hiányában (ui. az eddigi DNS vizsgálatok mindenesetre nem támasztották alá a dolgot, bár kétségtelen, hogy ez az eredmény önmagában nem cáfoló erejű – hiszen ha csak kismértékű keveredés volt a két populáció között, vagy esetleg csak keresekedtek, de nem keveredtek, annak mára aligha maradhatott nyoma), maradtak a közvetettebb dolgok, mint jellegzetes alakú kenuk, vagy éppen mindkét helyen előforduló, de csak az egyik helyen őshonos háziállatok ill. -növények. Utóbbira példa a batáta (vagy édesburgonya), amely ugyan az Újvilágból ered, de i.e. 1000 környékén már előfordult a Cook szigeteken, a régészeti leletek tanúsága szerint. Azonban ennél is érdekesebbnek bizonyult egy hétköznapi háziállat, a tyúk története.
A történelem folyamán a csirkét több helyen is háziasították, egyebek mellett Óceániában is. Vanatau és Tonga szigetén, az egykori emberi települések környékén, 3000 éves maradványai is előforulnak arra utalva, hogy a Lapita kultúra tagjai már felismerték a háziszárnyasok hasznát. A polinézek Melanéziából startoltak úgy ~3300 éve és az óceániai szigetvilág fokozatos belakását követően úgy ezer éve már Hawaii-ra is eljuttak, majd párszáz évvel később (de még bőven az európaiak előtt) a Húsvét szigeteket és Új Zélandot is megörvendeztették a jelenlétükkel (a moák mérsékelt örömére). Ha megnézzük a térképen, ezek egészen elképesztő távolságok – főleg a korabeli hajókkal -, ha ezeket sikerült abszolválni, akkor D-Amerika elérése teljesen benne van/volt a pakliban.
Olyannyira, hogy egy chilei ásatáson előkerült ~620 éves csirkecsontok mitokondriális DNS (mtDNS) analízise során kiderült, hogy azok teljesen megegyeznek Tongáról és Szamoáról előkerült még régebbi ill. hasonló korú maradaványok genetikai anyagával. Sőt, mind ez a szekvencia, mind a Chilében ma élő csirkékből kinyert mtDNS, sokkal jobban hasonlít a napjaink délkelet-ázsiai csirkéiben találhatóra, mint arra, amit európai rokonukból tudunk előbányászni. Márpedig ez azt jelenti – a képen látható Elizabeth Matisoo-Smith szerint -, hogy a polinéz csirkék leszármazottai voltak azok, amelyekkel pl. Pizarro az inkáknál találkozott, és még ma is ez a vérvonal kapirgál D-Amerika nyugati partjai mentén.
Storey AA, Ramirez JM, Quiroz D, Burley, DV, Addison, DJ et al. (2007) Radiocarbon and DNA evidence for a pre-Columbian introduction of Polynesian chickens to Chile. PNAS 104: 10335-10339.
 Szűk egy éve
Szűk egy éve  Ha már néhány napja egy ismert ökológiai szabályt igazoló megfigyeléről
Ha már néhány napja egy ismert ökológiai szabályt igazoló megfigyeléről  Ha véginézünk a
Ha véginézünk a  A szigetek élővilága mindig különleges, gondoljunk csak Új Zéland egészen egyedi madár faunájára vagy Galapagos pintyeire és
A szigetek élővilága mindig különleges, gondoljunk csak Új Zéland egészen egyedi madár faunájára vagy Galapagos pintyeire és 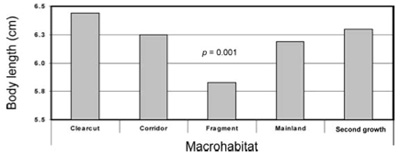 A Washington államban végzett kutatás végén a kutatók, több rágcsáló- (egér- ill. pocok-) fajt vizsgálva arra jutottak, hogy azok mérete az utóbbi mintegy három évtized alatt (ennyi a max. kora a vizsgált élőhely-töredékeknek) szignifikánsan csökkent más élőhelyen leledző rokonaikhoz képest. Mivel az egerek amúgy is igen sikeresek
A Washington államban végzett kutatás végén a kutatók, több rágcsáló- (egér- ill. pocok-) fajt vizsgálva arra jutottak, hogy azok mérete az utóbbi mintegy három évtized alatt (ennyi a max. kora a vizsgált élőhely-töredékeknek) szignifikánsan csökkent más élőhelyen leledző rokonaikhoz képest. Mivel az egerek amúgy is igen sikeresek 
 Az emberi beszédnek számos genetikai komponense van: a légcső beszédhez szükséges anatómiai jellegzetességeinek fejlődése (akárcsak bármely más szervünk) egyértelműen számos gén interakciójától függ, és nincs ez nagyon másképpen az agy beszédképzéshez ill. -értéshez való "drótozásával" sem.
Az emberi beszédnek számos genetikai komponense van: a légcső beszédhez szükséges anatómiai jellegzetességeinek fejlődése (akárcsak bármely más szervünk) egyértelműen számos gén interakciójától függ, és nincs ez nagyon másképpen az agy beszédképzéshez ill. -értéshez való "drótozásával" sem.