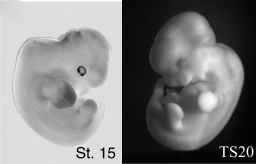
Az élővilág változatosságának legfőbb oka az embrionális fejlődés során bekövetkező kisebb-nagyobb változásokban rejlik. Általában minnél közelebbi rokon két faj, ezek a változások annál később válnak nyilvánvalóvá az egyedfejlődés során. Leggyakrabban nincs is szó új gének megjelenéséről, egyszerűen meglevő gének szabályozása változik meg egy kicsit. Mint a mellékelt ábrán látható pl. az embrionális fejlődés egészen késői szakaszáig a denevér fejlődése (bal oldalon) kísértetiesen hasonlít az egér fejlődésére (jobb oldalon). Már jól elkülönülnek az egyes testtájak, de még nyoma sincs a későbbi szárnynak és hegyes füleknek. Ezek később válnak láthatóvá, de a hozzájuk vezető molekuláris változások az ábrán látható idő tájékán következnek be.

A denevérek alakja nem új keletű, már az 50 millió éves fosszíliák (fent) a ma élő utódiakhoz (lent) hasonló szárnystruktúrával rendelkeztek. Ez pedig azt jelenti, hogy a hüvelykujjuk kivételével ujjperceik meghosszabodtak, s a köztük feszülő bőrredő révén úgynevezett „kéz-szárnyat” alkotnak. (Ezzel szemben a madarak szárnya ún. „kar-szárny” – amely a teljes kar módosulása révén alakult ki -, a Pterosaurusok szárnya pedig „ujj-szárny” volt – itt egyetlen ujj részeinek a meghosszabbodása képezte a szárny alapját.) Csak erre az alakra nézve, világossá válik, hogy a szárnykeletkezés kezdeti lépéseit valahol a mellső végatg fejlődése során kell keresni.
A végtagbimbók fejlődése és növekedése során fokozatosan alakulnak ki a kart alkotó csontok. Először sejtek vándorlása és gyülekezése révén egy porc alapú „váz” jön létre, amelyet később fokozatosan helyettesít a csontszövet. Ez az a pont ahol az egér és a denevér végtagjainak kialakulását figyelve egy fontos különbséget lehet észrevenni: utóbbiak esetében a kialakuló ujjak porcos váza nem áll meg a növekedésben.
 A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd egyes halak és pintyek arcberendezésének alakulását), de végtagok esetén a tudományos irodalom további tippekkel is szolgál. Ha egerek végtagbimbóit Bmp2 gazdag médiumban nevelgetjük: az ujjak hosszabra nőnek, ellenben, ha egy Bmp anatgonistát, Noggint keverünk a tápoldatba, az tompa, rövid ujjakhoz jutunk. Ezen a gondolati úton végighaladva nem nehéz kitalálni, mi is az első teendő, ha a denevér ujjak megnyúlásának okát keressük: megvizsgálni, mi történik a kézben kifejeződő bmp2 génnel.
A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd egyes halak és pintyek arcberendezésének alakulását), de végtagok esetén a tudományos irodalom további tippekkel is szolgál. Ha egerek végtagbimbóit Bmp2 gazdag médiumban nevelgetjük: az ujjak hosszabra nőnek, ellenben, ha egy Bmp anatgonistát, Noggint keverünk a tápoldatba, az tompa, rövid ujjakhoz jutunk. Ezen a gondolati úton végighaladva nem nehéz kitalálni, mi is az első teendő, ha a denevér ujjak megnyúlásának okát keressük: megvizsgálni, mi történik a kézben kifejeződő bmp2 génnel.
A fentiek ismeretében az eredmény nem teljesen meglepő: a denevérek fejlődő mellső végtagjának ujjaiban mind a bmp2 expresszió, mind a Bmp-k által szabályozott Smad jelátviteli út aktivitása sokszorosa annak, amit a hátsóvégtagjukban, vagy egerek mellső végtagjában figyelhetünk meg. Azaz a szárnykialakulás egy fontos (bár nem kizárólagos) tényezője a szóbanforgó gén aktivitásának felerősödése lehetett egy olyan helyen, ahol amúgy is kifejeződött az egyedfejlődés során.

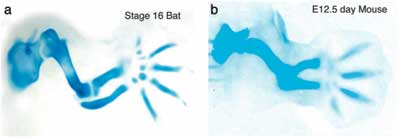
A denevérek mellső végtagja fejlődő ujjperceinek sejtjeiben (kék) lényegesen magassabb a Bmp fehérjék (piros) szintje, min a hátsó végatag ujjperceiben, vagy az egerek mellső végtagjában (e-g). Mindez a bmp2 gén magasabb aktivitásával függ össze (h) és ennek következményeként a Smad jelátviteli út is sokkal több sejtben kerül aktiválásra (piros) a denevér kéz ujjaiban mint lábukban vagy az egerekben (i-k).
Azok pedig akik azt szeretnék tudni, hogy a későbbiekben mi történik a denevér embrióval, a Pharyngulán elégíthetik ki kiváncsiságukat. 😉
Sears KE, Behringer RR, Rasweiler JJ, Niswander LA (2006) Development of bat flight: Morphologic and molecular evolution of bat wing digits. Proc. Nat. Acad. Sci. USA 103(17): 6581-6586.

 Nem igazán múlhat el hét valamilyen Hox gén sztori nélkül (max. az én hibámból ;-)), szóval lássuk, mi is keltett az elmúlt napokban nagy hírverést biológus berkekben (s mint a végén látni fogjuk, nemcsak ott).
Nem igazán múlhat el hét valamilyen Hox gén sztori nélkül (max. az én hibámból ;-)), szóval lássuk, mi is keltett az elmúlt napokban nagy hírverést biológus berkekben (s mint a végén látni fogjuk, nemcsak ott).
 Darwin pintyei mindmáig a természetes szelekció kinézet (fenotípus) formáló erejének talán legjobb szemléltetői maradtak. Ma már gimis anyag, hogy a Galapagos szigeteken fellelhető magok mérete miként formálta a pinty (Geospiza) fajok csőrének méretét és alakját, s mint az már egyszer
Darwin pintyei mindmáig a természetes szelekció kinézet (fenotípus) formáló erejének talán legjobb szemléltetői maradtak. Ma már gimis anyag, hogy a Galapagos szigeteken fellelhető magok mérete miként formálta a pinty (Geospiza) fajok csőrének méretét és alakját, s mint az már egyszer 
 A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.
A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.
 Humán immunológiát, vagy egyszerűen csak az
Humán immunológiát, vagy egyszerűen csak az  A blog régi olvasói talán még emlékeznek arra, hogy anno egy
A blog régi olvasói talán még emlékeznek arra, hogy anno egy