
 Nehéz ma már átélni a döbbenetet, amelyet a 18. század végén az európai felfedezők érezhettek, mikor is először szembesültek a kacsacsőrű emlőssel. Nyilván az sem könnyített helyzetükön, hogy Angliában sokan egyszerű szélhámosoknak tekintették őket, akik koruk "olajfaló" hoaxában mesterkednek – hiszen ki gondolhatná komolyan, hogy kacsa csőrrel és hártyás lábbal ellátott emlősök valóban léteznek. Nem ez lett volna az első eset, hasonló próbálkozásokra, így aztán a skót anatómus, Robert Knox gyanakvása igazán érthetőnek tűnik: "ennek a különleges állatnak az első példányai […] olyan hajókon keresztül jutottak Angliába, amelyek előzőleg az Indiai tenger vizeit szelték, s ez a tény önmagában is gyanakvást kellene keltsen a természettudósban, aki tisztában van azokkal a praktikákkal, amelyekkel tehetséges kínaiak verték át gyakorta a kalandvágyó európaiakat."
Nehéz ma már átélni a döbbenetet, amelyet a 18. század végén az európai felfedezők érezhettek, mikor is először szembesültek a kacsacsőrű emlőssel. Nyilván az sem könnyített helyzetükön, hogy Angliában sokan egyszerű szélhámosoknak tekintették őket, akik koruk "olajfaló" hoaxában mesterkednek – hiszen ki gondolhatná komolyan, hogy kacsa csőrrel és hártyás lábbal ellátott emlősök valóban léteznek. Nem ez lett volna az első eset, hasonló próbálkozásokra, így aztán a skót anatómus, Robert Knox gyanakvása igazán érthetőnek tűnik: "ennek a különleges állatnak az első példányai […] olyan hajókon keresztül jutottak Angliába, amelyek előzőleg az Indiai tenger vizeit szelték, s ez a tény önmagában is gyanakvást kellene keltsen a természettudósban, aki tisztában van azokkal a praktikákkal, amelyekkel tehetséges kínaiak verték át gyakorta a kalandvágyó európaiakat."
A különös lény azonban csak nem kart eltűnni, újabb és újabb felfedezők és kutatók számoltak be róla, míg aztán végül kétségtelenné vált, hogy a kacsacsőrű emlős valóban létezik. Különcsége miatt természetesen élénk érdeklődésre tartott számot, aminek azonban az élőhelyeinek messzisége szabott igencsak szűkös határt. Így történhetett, hogy a felfedezéstől számítva majdnem száz évnek kellett eltelnie, hogy egyértelművé váljon, az Ornithorhyncus anatinus (amint latinul végül elnevezték), tojással szaporodik.
Az evolúciós elmélet térnyerésével az is nyilvánvalóvá vált, hogy a viszonylag kisszámú, tojással szaporodó, kloakás emlős faj egyfajta "élő kövületnek" is tekinthető, hiszen az emlősök evolúciójának egy olyan lépését dokumentálják, amikor az emlők és a tejtermelés kialakult már, de a méhlepény még váratott magára. Persze ez nem azt jelenti, hogy a kacsacsőrűek (ill. más kloakások) felett megállt az idő; továbbhaladtak ők is saját evolúciós ösvényükön, magyarán a maguk módján ők is sokat változtak. Ez utóbbiról tanúskodik a napokban elkészült genomszekvenciájuk is.
A száraz tények mellett (az 52 kromoszómából álló genom durván 18,000 gént tartalmaz, és kb. fele ismétlődő szekvenciákból áll), természetesen ott figyelnek azok a nyalánkságok is, amelyek egyenként is külön posztokat érnek. (Pontosabban értek, hiszen sokról – pl. sajátos opszin génjeikről, vagy a szikfehérjéikről – már írtam.) Az egyik, az az állatvilágban egészen egzotikus genetikai nemmeghatározási mechanizmus, amely nem kevesebb mint öt (!!) pár kromoszómát igényel. Vagy ott van, az emlősök közül szintén csak kloakásokra jellemző elektrorecepció (a vízben élő kacsacsőrű emlős szorosan zárt szemekkel, füllel és orral vadászik, csak erre a különleges érzékére hagyatkozva), illetve a hím kacsacsőrűek hátsó lábán levő méregmirigye.
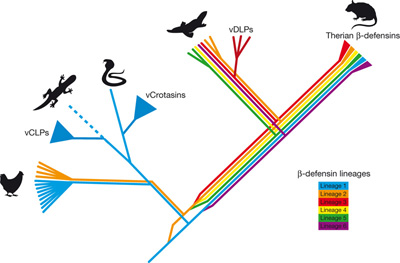 Mivel összességében is tucatnyinál kevesebb "mérgező" emlősről tudunk, ez a bájos kis vonás önmagában is vonzaná a kutatói tekinteteket. És nem is hiába: mint kiderült, a kacsacsőrű emlős és a különböző hüllő fajok méreganyagai, a konvergens evolúció eklatáns példájaként, azonos tőről fakadnak. Mindkét esetben, az egyébként az immunrendszerben fontos, beta-defensin gének duplikációjából eredeztethetőek a későbbi toxikus anyagok, bár hangsúlyozandó (mint azt a mellékelt ábra is tükrözi), a beta-defensinek duplikációja nem feltétlenül jelenti méreganyagok megjelenését. Az alaptézisen azonban ez sem változtat: még az olyan előzmény nélkülinek és drámainak tűnő evolúciós újítások, mint a méregkiválasztás, sem a semmiből keletkeznek, illetve egy jól alkalmazható receptre (kisebb-nagyobb változtatásokkal) néha többször is "rálel" a természetes szelekció.
Mivel összességében is tucatnyinál kevesebb "mérgező" emlősről tudunk, ez a bájos kis vonás önmagában is vonzaná a kutatói tekinteteket. És nem is hiába: mint kiderült, a kacsacsőrű emlős és a különböző hüllő fajok méreganyagai, a konvergens evolúció eklatáns példájaként, azonos tőről fakadnak. Mindkét esetben, az egyébként az immunrendszerben fontos, beta-defensin gének duplikációjából eredeztethetőek a későbbi toxikus anyagok, bár hangsúlyozandó (mint azt a mellékelt ábra is tükrözi), a beta-defensinek duplikációja nem feltétlenül jelenti méreganyagok megjelenését. Az alaptézisen azonban ez sem változtat: még az olyan előzmény nélkülinek és drámainak tűnő evolúciós újítások, mint a méregkiválasztás, sem a semmiből keletkeznek, illetve egy jól alkalmazható receptre (kisebb-nagyobb változtatásokkal) néha többször is "rálel" a természetes szelekció.
Warren WC, Hillier LW, Marshall Graves JA, Birney E, Ponting CP, et al. (2008) Genome analysis of the platypus reveals unique signatures of evolution. Nature 453: 175-183.
Hall BK (1999) The Paradoxical Platypus BioScience 49(3): 211-218.
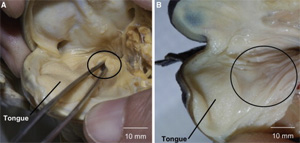 "A kétéltűek az első szárazföldi gerincesek, tüdővel lélegeznek", olvashatjuk
"A kétéltűek az első szárazföldi gerincesek, tüdővel lélegeznek", olvashatjuk  Bevallottan ignoráns ember révén, aki képes hónapokat leélni az ingyenes (pletyka)lapok böngészése nélkül, némi fáziskéséssel, csak most értesültem
Bevallottan ignoráns ember révén, aki képes hónapokat leélni az ingyenes (pletyka)lapok böngészése nélkül, némi fáziskéséssel, csak most értesültem 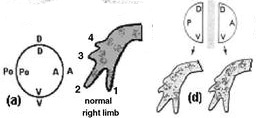
 Harminchat évvel ezelőtt, a horvát Adria egy zsebkendőnyi kis szigete, Pod Mr
Harminchat évvel ezelőtt, a horvát Adria egy zsebkendőnyi kis szigete, Pod Mr aru, sajátos kísérlet színhelyévé vált. A szomszédos, leheletnyit nagyobb, Pod Kopi
aru, sajátos kísérlet színhelyévé vált. A szomszédos, leheletnyit nagyobb, Pod Kopi te szigetéről öt pár olasz faligyíkot (Podarcis sicula) telepítettek át, hogy megnézzék … mi is történik.
te szigetéről öt pár olasz faligyíkot (Podarcis sicula) telepítettek át, hogy megnézzék … mi is történik.
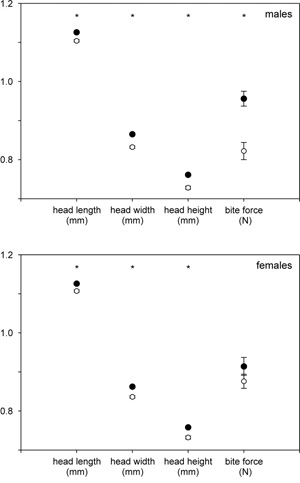 Visszatérve a gyíkokra, talán nem meglepő, ha itt is drámai változásoknak lehetünk tanúi, bár alig 30 generációnyi idő telt el. Külsőleg legszembetűnőbb, hogy a Pod Mr
Visszatérve a gyíkokra, talán nem meglepő, ha itt is drámai változásoknak lehetünk tanúi, bár alig 30 generációnyi idő telt el. Külsőleg legszembetűnőbb, hogy a Pod Mr Vannak olyan cikkek, ahol az ember csak vakarni tudja a fejét, hogy mit is mondjon újat. Mert minden
Vannak olyan cikkek, ahol az ember csak vakarni tudja a fejét, hogy mit is mondjon újat. Mert minden 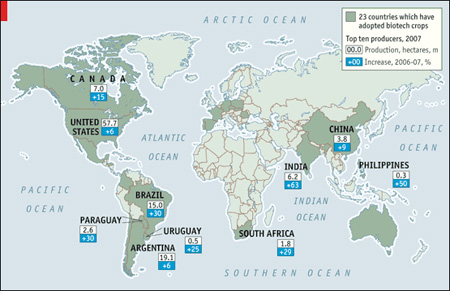 A GMO-s viták, és egyáltalán a GMO-val kapcsolatos attitűdök, egyik döntő problémája az az információs aszimetria, ami az elmúlt években a technológia pozitív és negatív vonatkozásaival kapcsolatban alakult ki. Míg a káros oldalakról (egyébként helyesen) rendszeresen beszámolnak a különböző médiumok, a pozitív hírek ritkán, vagy
A GMO-s viták, és egyáltalán a GMO-val kapcsolatos attitűdök, egyik döntő problémája az az információs aszimetria, ami az elmúlt években a technológia pozitív és negatív vonatkozásaival kapcsolatban alakult ki. Míg a káros oldalakról (egyébként helyesen) rendszeresen beszámolnak a különböző médiumok, a pozitív hírek ritkán, vagy  A bordás- (vagy fésűs) medúzákat, vagyis a
A bordás- (vagy fésűs) medúzákat, vagyis a 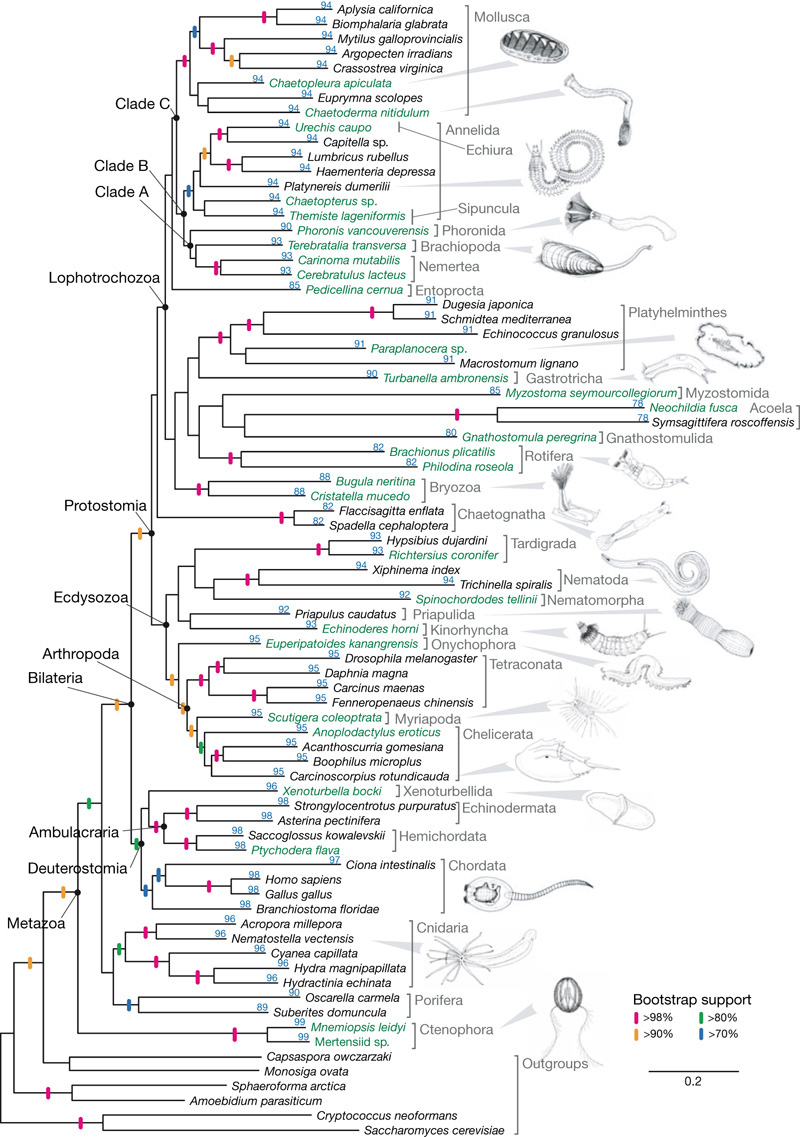
 Hagyományainkhoz híven (hiszen
Hagyományainkhoz híven (hiszen