
 Ha, túllépve a kreacionista álkifogásokon, valódi, tudományos vitára vagyunk kíváncsiak evolucionista berkekben, aligha találhatnánk fontosabbat, mint a "gradualizmus" és a "pontozott egyensúly" hívei közt zajlót.
Ha, túllépve a kreacionista álkifogásokon, valódi, tudományos vitára vagyunk kíváncsiak evolucionista berkekben, aligha találhatnánk fontosabbat, mint a "gradualizmus" és a "pontozott egyensúly" hívei közt zajlót.
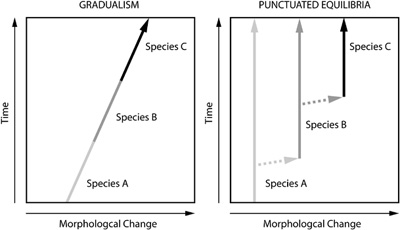
A két nézet közül előbbi a prominensebb és évtizedeken át egyeduralkodónak számított. Röviden, a klasszikus Darwini nézeteknek megfelelően, azt mondja ki, hogy az élővilágban a változások folyamatosan jelennek meg, és a változások felhalmozódása fokozatosan új fajok kialakulásához vezet.
A rivális elmélet Niles Eldredge (az American Museum of Natural History egyik jelenlegi kurátora) doktori téziséből született és Eldredge mellett a másik nagy propagálója kollegája, Stephen Jay Gould volt. Eldredge trilobita fosszíliák tanulmányozása közben arra figyelt fel, hogy ezek a fajok kinézetre több tízezer-, akár egy millió éven keresztül semmit sem változnak, majd hirtelen alakulnak át új fajokká. Gouldék a változatlan időszakot "sztázisnak" nevezték el, és amelett érveltek, hogy az evolúció nem az összes populációban bekövetkező, folyamatos változásban nyilvánul meg, hanem izolált közösségekben történő, (viszonylag) gyors váltásokban.
A vitának tengernyi irodalma van, de itt két kérdésre szűkíteném le a dolgot: létezik-e valóban sztázis, ill. a két elmélet valóban kizárja-e egymást?
Sztázis alatt a pontozott egyensúly hívei pangást, változatlanságot értenek, de nem akármilyen értelemben, hanem konkrétan morfológiai szempontból. (Mást nehezen lehet egy-egy trilbota fosszíliában tanulmányozni, így ez nem is meglepő.) Mivel a genetikai és morfológiai változások gyakran kéz a kézben járnak, ebből könnyedén következtethetnénk arra, hogy akkor ez azt is jelenti, hogy a sztázisban levő fajok örökítőanyaga sem nagyon változik. És nagyot hibáznánk.
 Egy "élő fosszílián" végzett genetikai vizsgálat – ha igaznak bizonyul – ezt a logikát ugyanis tételesen cáfolja meg.
Egy "élő fosszílián" végzett genetikai vizsgálat – ha igaznak bizonyul – ezt a logikát ugyanis tételesen cáfolja meg.
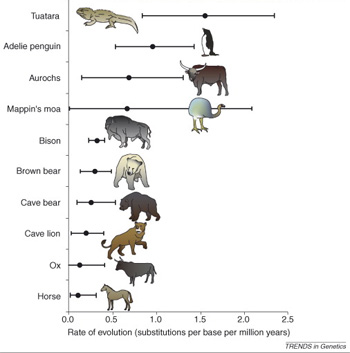
Az Új Zélandon élő tuatara, vagy hidasgyík, a hüllők egyik különös rendjének egyetlen ma is élő képviselője. Amennyire a fosszíliák alapján meg lehet ítélni, külalakra nem sokat változott az elmúlt évmilliók során, így aztán persze egész jó kis tanulmányi alanya lehet evolúciós kutatásoknak. Harminchárom ősi és negyvenegy jelenleg is élő tuatara mitokondriális DNS-t összevetve a kutatók arra a meglepő következtetésre jutottak, hogy a genetikai változások sokkal gyakoribbak ezekben a gyíkokban, mint pl. a legtöbb ma élő emlősfajban (amelyek közül sok, kifejezetten szembetűnő változáson ment át az utóbbi évmilliók során). Nyilván lehet, hogy egyszerűen az ősi DNS degradálódott annyira, hogy ma már csak sok hibával lehet rekonstruálni, de a kutatók mindeféle statisztikai ellenőrzésekkel (plusz a nagy mintaszámmal) védekeztek ez ellen, és a különbség elég nagy ahhoz, hogy ha néhány hibán nem is akadt meg a szemük, valósnak tűnjön.
Márpedig ha így van, az erősen abba az irányba mutatna, hogy a morfológiai sztázis nem zárja ki a genetikai gradualizmust, sőt. Elképzelhető, hogy azért észlelhetünk gyors morfológiai átmeneteket időnkéntaz evolúció során, mert a DNS szintjén, a mutációk már "előkészítették a terepet", hogy egy végső genetikai változás új kinézetet kölcsönözzön hordozójának.
Azt persze hangsúlyoznom kell, hogy még ha így is van, az nem jelenti azt, hogy a gradualizmus abban a formában igaz lenne, hogy a folyamatos változások mindig adott ütemben jelennének meg. Minden valószínűség szerint vannak gyorsabb ill. lassabb változást előidéző körülmények. De hogy tökéletes sztázis is létezne (genetikai értelemben) az erősen megkérdőjelezhető.
Eldredge N (2008) The Early "Evolution" of "Punctuated Equilibria". Evolution: Education and Outreach 1(2): 107-113. doi: 10.1007/s12052-008-0032-0
Hay JM, Subramanian S, Millar CD, Mohandesan E, Lambert DM (2008) Rapid molecular evolution in a living fossil. Trends Genet 24(3): 106-109.
 Pontosítás végett nem fosszíliákban levő génekről lesz szó (bár az is
Pontosítás végett nem fosszíliákban levő génekről lesz szó (bár az is  Amikor pár hete a Human Genome Diversity Project apropóján
Amikor pár hete a Human Genome Diversity Project apropóján 
 A véralvadás hasznos és fontos dolog, mint azt általában a vérzékenységben szenvedők napi problémái is jól szemléltetik. Egészen dióhéjban arról van szó, hogy a sérülés után (első lépésben) az érfalon keletkezett sérülést kell a szervezetünk valamiképpen betoldozza-foldozza, hogy meggátolja az ereinkben levő sejtes ill. nem-sejtes elemek kifolyását.
A véralvadás hasznos és fontos dolog, mint azt általában a vérzékenységben szenvedők napi problémái is jól szemléltetik. Egészen dióhéjban arról van szó, hogy a sérülés után (első lépésben) az érfalon keletkezett sérülést kell a szervezetünk valamiképpen betoldozza-foldozza, hogy meggátolja az ereinkben levő sejtes ill. nem-sejtes elemek kifolyását.
 Ha manapság környezettudatos ismerőseink arcára őszinte megdöbbenést, netán felháborodást szeretnénk varázsolni, akkor elég kedvesen megérdeklődni, hogy mutáns növényt, vagy egyenesen, lyesszasz, transzgénikus növényt esznek-e/ennének-e? A válasz többnyire "nem" lesz, gyakran kisebb beszéddel megfejeleve, amelyben az említett növények káros hatásait ecsetelik buzgó szorgalommal.
Ha manapság környezettudatos ismerőseink arcára őszinte megdöbbenést, netán felháborodást szeretnénk varázsolni, akkor elég kedvesen megérdeklődni, hogy mutáns növényt, vagy egyenesen, lyesszasz, transzgénikus növényt esznek-e/ennének-e? A válasz többnyire "nem" lesz, gyakran kisebb beszéddel megfejeleve, amelyben az említett növények káros hatásait ecsetelik buzgó szorgalommal.
 S hogy gyorsan példával is alátámasszam, mit is értettem
S hogy gyorsan példával is alátámasszam, mit is értettem 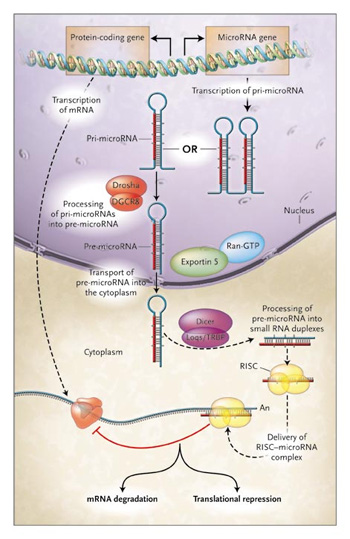 Bő tíz éve szinte véletlen megfigyelésekből indult a dolog és mára a biológia egyik legizgalmasabb területévé vált az RNS-függő génexpresszió-szabályozás tanulmányozása. Felfedezőinek máris
Bő tíz éve szinte véletlen megfigyelésekből indult a dolog és mára a biológia egyik legizgalmasabb területévé vált az RNS-függő génexpresszió-szabályozás tanulmányozása. Felfedezőinek máris