
 A bordás- (vagy fésűs) medúzákat, vagyis a Ctenophora törzs képviselőit, mint azt a nevük is mutatja, hosszabb ideje többé kevésbé a medúza- és polipéletformával egyaránt rendelkező csalánozókkal (Cnidaria) vették egy kalap alá.
A bordás- (vagy fésűs) medúzákat, vagyis a Ctenophora törzs képviselőit, mint azt a nevük is mutatja, hosszabb ideje többé kevésbé a medúza- és polipéletformával egyaránt rendelkező csalánozókkal (Cnidaria) vették egy kalap alá.
Pedig a hasonlóság tényleg leginkább csak felületetes: mindkét csoport tagjait áttetsző, látszólag körkörös szimmetriájú test jellemzi, na és a standard három csíralemezből is csak kettő (endo- és ectoderma) lelhető fel minden kétséget kizárólag. A kulcsjegynek számító csalánsejtek (amiről a csalánozók a nevüket is kapták) viszont a bordásmedúzákból hiányoznak, helyettük ragadós, ún. colloblaszt sejtekkel ragadják meg mikroszkópikus prédáikat.
Mivel a ctenophorák testében valódi izmok is felfedezhetőek (ami miatt jópár kutató szerint valójában itt kialakul a harmadik csíralemez, a mezoderma is, amelyből a legtöbb „felsőbbrendű” állatban az izmok származnak), sokak hajlamosak (voltak) a kétoldali szimmetriával rendelkező állatokhoz (Bilateria) közelebb sorolni őket. Ezzel viszont az legnagyobb probléma, hogy már a korai – és hiányos – molekuláris rendszerezési adatok sem támasztották alá.
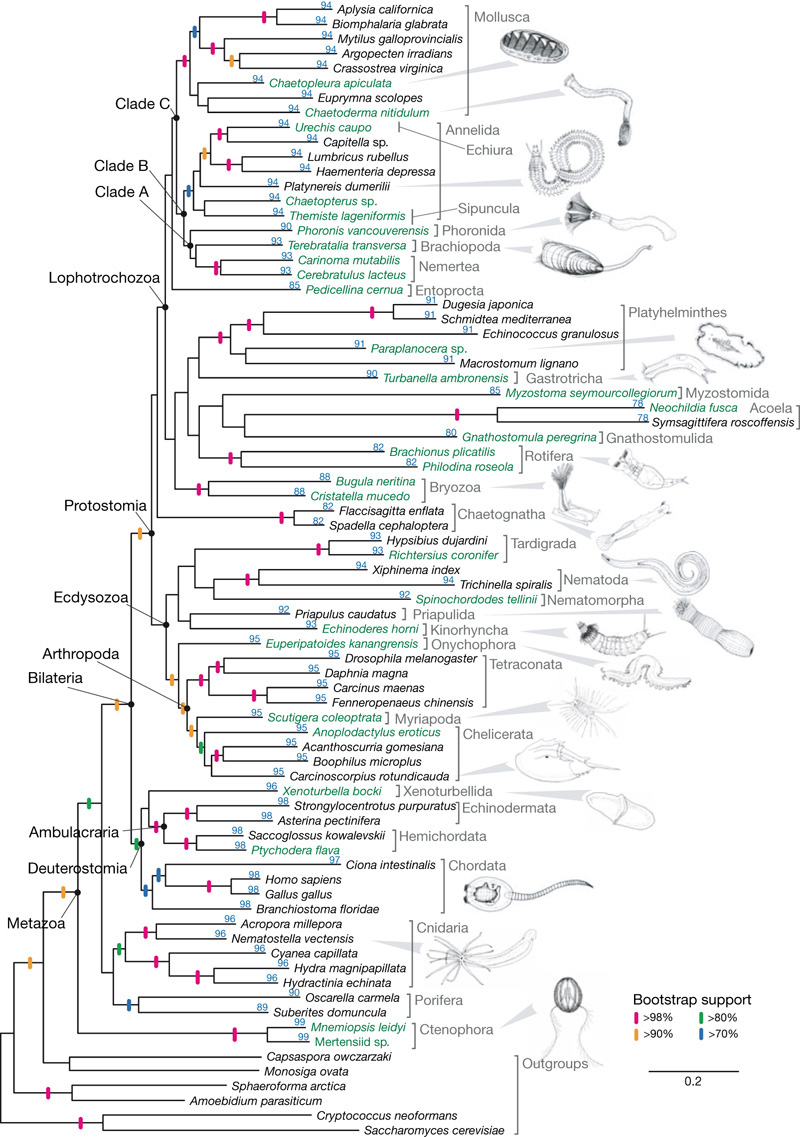
Szóval ilyen és ehhez hasonló vitatottabb rendszertani problémák megoldására a közelmúltban néhány kutató nekiült egy minden eddiginél részletesebb rendszertani fát gründoljon genomi DNS szekvenciák alapján (klikk az alábbi ábrára). Huszonegy állattörzs képviselőit vették górcső alá (ebből tizenegy (!!) eddig nem szerepelt ilyen típusú vizsgálatban), és masszív, majd negyven megabázisnyi szekvenciát használtak. Az eredmények sok szempontból érdekesek, bár legtöbb esetben ma már nem teljesen váratlanok. Egyrészt igazolást nyert, hogy a klasszikus testüreg kialakulása szerinti felosztás (Acoelomata, Pseudocoelomata, Eucoelomata) teljesen tarthatatlan. Helyette a leendő végbélnyílás eredete szerint megkülönböztethetünk ó- és újszájú állatokat (Protostomia és Deuterostomia), illetve előbbiek közt az igazán nagy különbség a lárvák típusa szerint van. Létezik az ún. Ecdysozoa csoport, amelybe az életük során vedlő fajok, pl. rovarok, rákok, pókok tartoznak, és a Lophotrochozoa csoport, amelybe mindefajta férgek és puhatestűek (csigák, oktopuszok, kagylók) esnek.
Bizonyossá vált továbbá az is, hogy a korábbi morfológiai bélyegeken alapuló fákkal ellentétben, az előgerinchúrosok közelebbi (bár sokban módosult) rokonai a gerinceseknek, mint a fejgerinchúrosok.
A legérdekesebb eredmény (értelemszerűen ;-)) a bordásmedúzákhoz kapcsolódik: az új fa szerint ezek még a szivacsoknál is távolabbi unokatestvérei minden más állatnak! Ha ez valóban így van (és még néhány megabázisnyi szekvencia, plusz négy-öt újabb faj vizsgálata ezt hamarosan eldöntheti), az jópár fogas kérdést vet fel, hiszen a bordásmedúzák testfelépítése komplexebb, mint a szivacsoké, amelyekből valódi szövetek is hiányoznak. Lehetséges, hogy a szivacsok „primitívsége” ugyanúgy másodlagos redukció eredménye, mint a parazita férgeké? Vagy a bordásmedúzák a többi állattól némileg függetlenül hozták létre a saját testfelépítésüket? (Vagy, és azért még ne zárjuk ki, a ctenophorák genetikai anyaga gyors evolúción ment át és ez becsapta az ilyesmire egyébként ma már felkészített, filogenetikai fakészítő algoritmusokat?) Pontosabb képet csak úgy nyerhetünk, ha jobban megismerjük ezeknek a különleges lényeknek a biológiáját, így koránt sem lennék meglepődve, ha a következő hónapokban jópár labor sok energiát fektetne a bordásmedúzák fejlődési nüanszainak feltárásába.
(A bordásmedúza fényképe a JelliesZone-ról származik.)
Martindale MQ, Henry JQ. (1999) Intracellular fate mapping in a basal metazoan, the ctenophore Mnemiopsis leidyi, reveals the origins of mesoderm and the existence of indeterminate cell lineages. Dev Biol 214(2): 243-257.
Dunn CW, Hejnol A, Matus DQ, Pang K, Browne WE, et al. (2008) Broad phylogenomic sampling improves resolution of the animal tree of life. Nature 452: 745-749.
Juj de jó lett ez a poszt!