
 Jó pár évvel ezelőtt, a blog hajnalán, az egyik legizgalmasabb és legfelkapottabb kérdés a biológiában az volt, hogy a különböző fajok (alfajok) morfológiai sajátosságai miképpen alakulnak ki a fejlődés során. Az evodevo tudománya – mert arról van szó – azóta kicsit kiment a mainstream divatból, de ha engem kérdezne bárki, szerintem továbbra is az egyik legizgalmasabb kérdés, amivel biológus foglalkozhat.
Jó pár évvel ezelőtt, a blog hajnalán, az egyik legizgalmasabb és legfelkapottabb kérdés a biológiában az volt, hogy a különböző fajok (alfajok) morfológiai sajátosságai miképpen alakulnak ki a fejlődés során. Az evodevo tudománya – mert arról van szó – azóta kicsit kiment a mainstream divatból, de ha engem kérdezne bárki, szerintem továbbra is az egyik legizgalmasabb kérdés, amivel biológus foglalkozhat.
A terület egyik úttörője, Sean B. Carroll mellett David Kingsley volt, aki azóta is azzal a nagyon egyszerű paradigmával foglalkozik, hogy milyen genetikai különbségek találhatók a háromtüskés pikó páncélos, tüskés tengeri és páncéltalan, tüskétlen édesvízi formái között.
Ahogy azt már korábban leírtam, ezzel a megközelítéssel leltek arra a pitx1 szabályozó mutációra, ami a tüskék eltűnését eredményezi, illetve így térképezték fel csontlemezek redukciójáért felelős ectodysplasin (eda) gént érintő mutációt is.
 A barlangi élet különös alkalmazkodásokat követel meg azoktól az élőlényektől, akik a létnek ezt a sötét formáját választják. A jóformán örök sötétségben fölöslegessé válik a felszínen oly fontos látás, lényegtelenné a bármilyenfajta mintázat, más érzékek és más jellegek válnak fontossá az életbenmaradás és fajfenntartás céljából.
A barlangi élet különös alkalmazkodásokat követel meg azoktól az élőlényektől, akik a létnek ezt a sötét formáját választják. A jóformán örök sötétségben fölöslegessé válik a felszínen oly fontos látás, lényegtelenné a bármilyenfajta mintázat, más érzékek és más jellegek válnak fontossá az életbenmaradás és fajfenntartás céljából.  A lóháziasítás egyik eredménye a különböző fajták minta-diverzitása, ami ha nem is annyira látványos, mint mondjuk kutyák esetében, de mindenképpen figyelemre méltó.
A lóháziasítás egyik eredménye a különböző fajták minta-diverzitása, ami ha nem is annyira látványos, mint mondjuk kutyák esetében, de mindenképpen figyelemre méltó.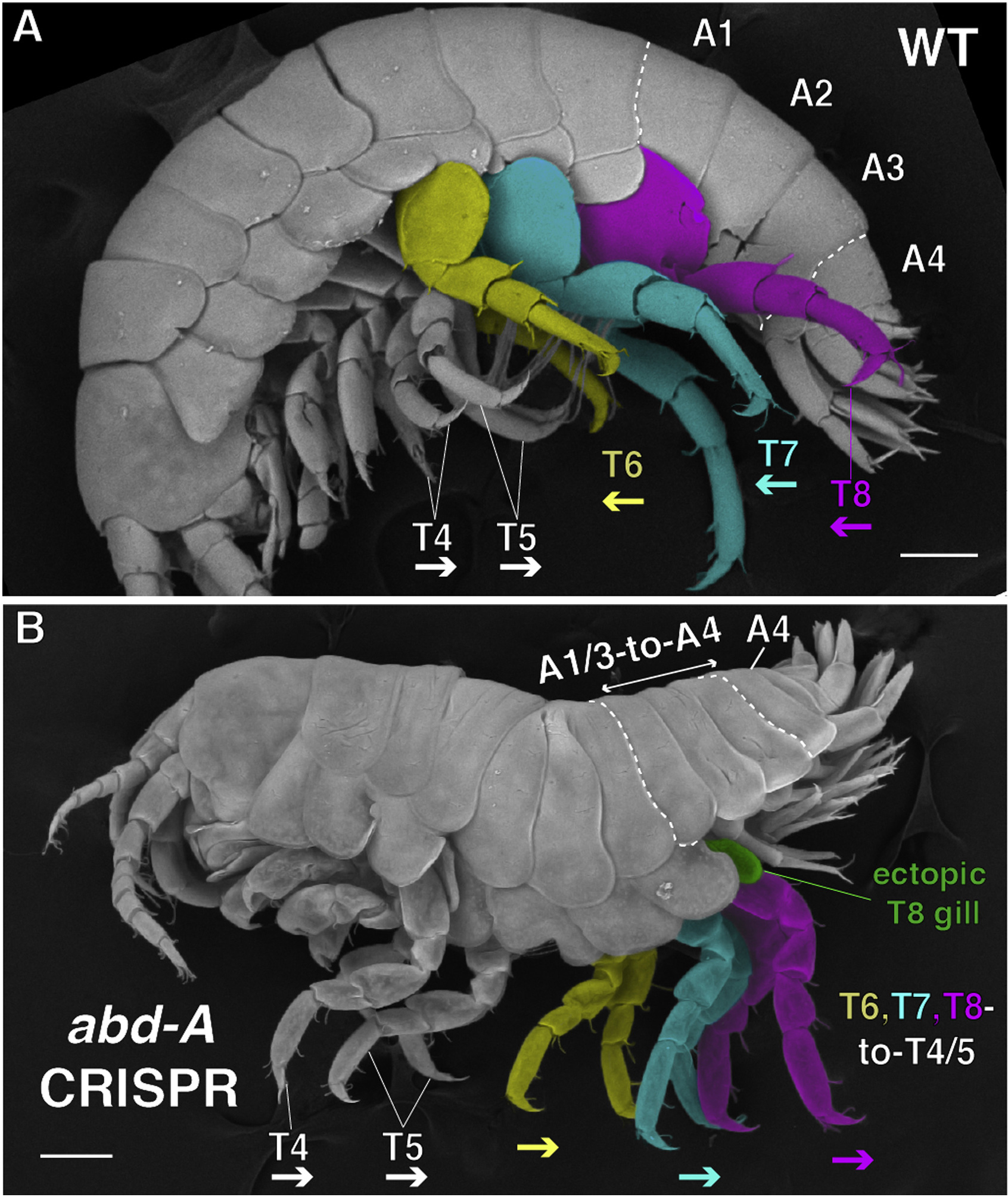 Vannak témák, amelyek végigkövetik a blog most már évtizedes történetét, és az egyik legrégebbi történet, aminek szálait göngyölítgetjük, arról szól, hogy az élővilág evolúciója során miként alakult ki az élőlények formagazdagsága.
Vannak témák, amelyek végigkövetik a blog most már évtizedes történetét, és az egyik legrégebbi történet, aminek szálait göngyölítgetjük, arról szól, hogy az élővilág evolúciója során miként alakult ki az élőlények formagazdagsága.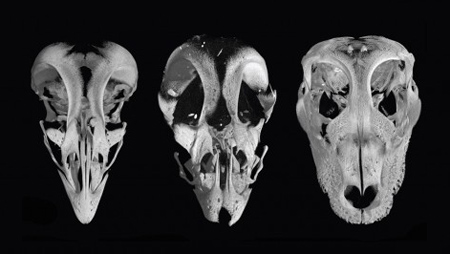 A Harvardon dolgozó
A Harvardon dolgozó  Amikor anno a lándzsahal, vagyis
Amikor anno a lándzsahal, vagyis 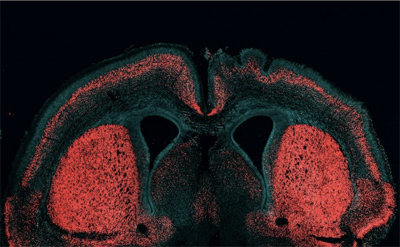 Az
Az  Szinte napra pontosan Darwin
Szinte napra pontosan Darwin 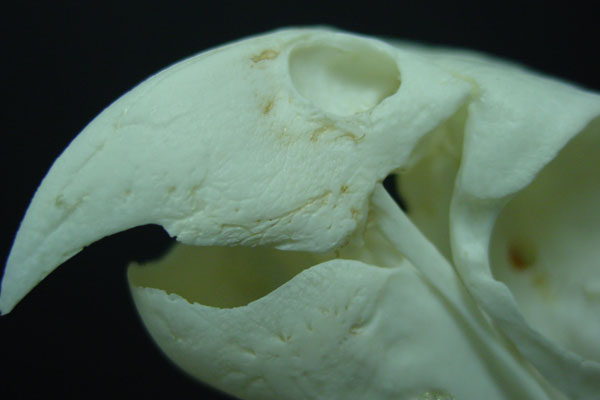 A madarak törzsfája mindig is bonyolultnak tűnt, nem véletlenül volt szükséges 48 különböző faj genomjának megszekvenálásához, hogy végre
A madarak törzsfája mindig is bonyolultnak tűnt, nem véletlenül volt szükséges 48 különböző faj genomjának megszekvenálásához, hogy végre 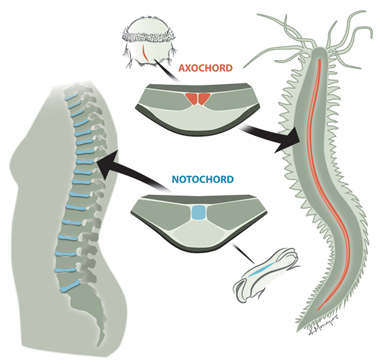 Szélesebb rendszertani csoportunk közös jellemzője, hogy minden idetartozó faj a fejlődés valamelyik stádiumában egy jellegzetes képletet növeszt, a gerinchúrt.
Szélesebb rendszertani csoportunk közös jellemzője, hogy minden idetartozó faj a fejlődés valamelyik stádiumában egy jellegzetes képletet növeszt, a gerinchúrt.