
Ahogy egy-egy szó jelentését is nagymértékben befolyásolja a szövegkörnyezet, úgy az egyes fehérjék „jelentését”, funkcióját is nagymértékben befolyásolja, hogy milyen molekuláris milliőbe kerülnek, milyen más fehérjékkel kerülnek kölcsönhatásba.
A BMP fehérjék visszatérő vendégeink, és már korábban többször is szóltam arról, hogy az egyik legkarakterisztikusabb tulajdonságuk, hogy elősegítik a csontnövekedést – pl. a denevérszárny ujjperceinek meghosszabbodását is. De a BMP-knek tucatnyi más feladatuk is van az egyedfejlődés során, s ezek gyakran térben és időben szinte átfednek a korábban már említett feladatokkal (a funkcionális különbség a molekuláris partnerek másságából adódik).
Ennek egészen jó példája a denevérszárnyak esete (hogy témánál maradjunk), ahol az ujjpercek porcosodását és megnyúlását megelőzően a bmp expresszió pont az ujjak közötti szövetekben figyelhető meg, ahol a szóbanforgó sejtek elpusztulását idézi elő. Ezért nem lesz aztán semmilyen hártya az ujjaink között. Mármint nekünk, mert más fajok esetében van erre példa. Pl. a kacsák esetében, ahol elég pontos képünk van arról, hogy miként gátolódik a BMP-függő sejthalál az ujjközti területeken. Egyrészt a BMP jelátvitel egyik effektora, az msx gén nem fejeződik ki az említett régióban (ellentétben pl. a csirkékkel, vagy egerekkel), másrészt pedig megjelenik a gremlin nevű gén terméke, amely egy igen potens BMP anatgonista.
Most néhányan annak eredtek a nyomába, hogy mennyire hasonló okok miatt marad meg a hártya a denevér ujjai között. Mint kiderült, még a konvergens evolúció sem lép kétszer tökéletesen ugyanabba a folyóba: az msx expresszió a denevér embrióban gyakorlatilag megegyezik az egérembrióban megfigyeltekkel (vagyis ez nem lehet az oka az apoptózis elmaradásának). Viszont a gremlin expresszió valóban megjelenik a denevérek mellső végtagkezdeményében. S bár ez már önmagában is elegendő lehetne a sejtpusztulás elmaradásának magyarázatra, a jelek szerint működik még egy párhuzamos mechanizmus is: egy fibroblast growth factor (fgf) gén is elkezd kifejeződni a leendő hártyában, ami szintén a sejthalál gátlásához vezet.
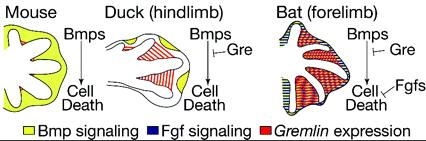
Summa-summarum, a hártyás szárny (vagy éppen láb) nem a semmiből jelent meg az evolúció során; mindig is jelen volt a négylábú gerinces embriók fejlődésének egy adott szakaszában. S amikor az élőlénynek haszna számazhatott abból, ha ez a hártya megmarad, a szelekció révén kialakultak azok a molekuláris „fékek”, amelyek meggátolták elhalását.
Weatherbee, SD, Behringer, RR, Rasweiler, JJ, Niswander, LA (2006) Interdigital webbing retention in bat wings illustrates genetic changes underlying amniote limb diversification. PNAS 103: 15103-15107.

 A kelet afrikai Nagytavakban (Malawi, Viktória és Tanganyika) élő sügérfajok mindig is az evolúció kirakatállatainak számítottak. A tavakban ma megfigyelhető több száz bölcsőszájú sügérfaj (cichlid) egyetlen közös ősből alakult ki, de mára a tavak biztosította összes élőhelyet kihasználják. Algaevő aljzatlakók és plaktonevő felszíni fajok színes kavalkádja jött létre élővilág egyik legnagyobb adaptív radiációja során.
A kelet afrikai Nagytavakban (Malawi, Viktória és Tanganyika) élő sügérfajok mindig is az evolúció kirakatállatainak számítottak. A tavakban ma megfigyelhető több száz bölcsőszájú sügérfaj (cichlid) egyetlen közös ősből alakult ki, de mára a tavak biztosította összes élőhelyet kihasználják. Algaevő aljzatlakók és plaktonevő felszíni fajok színes kavalkádja jött létre élővilág egyik legnagyobb adaptív radiációja során. A szépségnek ára van, még akkor is, ha bogarakról van szó. Pár hete már
A szépségnek ára van, még akkor is, ha bogarakról van szó. Pár hete már  Egy-egy bálnacsontvázzal szembesülve az átlagember első (és gyakran egyetlen) gondolata, hogy „hát igen, ez valóban bazi nagy”. Azonban a fizikai nagyság el- és felismerése mellett gyakran hajlamosak vagyunk elsuhanni az apróbb részletek mellett (persze néha kontár preparátorok is segítenek ebben).
Egy-egy bálnacsontvázzal szembesülve az átlagember első (és gyakran egyetlen) gondolata, hogy „hát igen, ez valóban bazi nagy”. Azonban a fizikai nagyság el- és felismerése mellett gyakran hajlamosak vagyunk elsuhanni az apróbb részletek mellett (persze néha kontár preparátorok is segítenek ebben). Állati szarvakról beszélve (már csak nyelvi okok miatt is) elsősorban mindenkinek a szarvas szokott beugrani, pedig az igazi nagymesterei a szarvnövesztésnek a bogarak között találhatók. A
Állati szarvakról beszélve (már csak nyelvi okok miatt is) elsősorban mindenkinek a szarvas szokott beugrani, pedig az igazi nagymesterei a szarvnövesztésnek a bogarak között találhatók. A 
 Darwin pintyei mindmáig a természetes szelekció kinézet (fenotípus) formáló erejének talán legjobb szemléltetői maradtak. Ma már gimis anyag, hogy a Galapagos szigeteken fellelhető magok mérete miként formálta a pinty (Geospiza) fajok csőrének méretét és alakját, s mint az már egyszer
Darwin pintyei mindmáig a természetes szelekció kinézet (fenotípus) formáló erejének talán legjobb szemléltetői maradtak. Ma már gimis anyag, hogy a Galapagos szigeteken fellelhető magok mérete miként formálta a pinty (Geospiza) fajok csőrének méretét és alakját, s mint az már egyszer 
 A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.
A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.

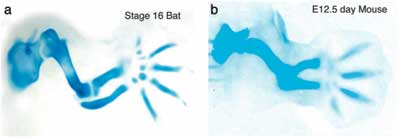 A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd
A kérdés tehát adott: milyen gének termékei a lehetnek felelősek ezért? A válasz pedig nem is annyira nehéz. Van ugyanis egy fehérje család, amely már a nevébő adódóan is jó jelölt ezek az ún. Bmp-k azaz Bone Morphogenetic Proteinek. A név (persze) nem véletlen számos olyan esetről tudunk, amikor különböző Bmp-k révén jellegzetes változások következtek be a csontozat kialakulásában. Néhányról korábban már én is említést tettem (lásd 