
 Mint azt minden természetjáró tapasztalhatta, a magashegyi környezet alacsony oxigéntartalma elég megterhelő a szervezet számára. Mivel a vér nem tud elegendő oxigént szállítani az izmokhoz, különösen az edzetlen túrázó a legkisebb megterheléstől is lihegni kezd, a legenyhébb szellőtől is elfárad. A probléma kiküszöbölésének egyik útja a rendszeres légzésszám megnövelése (ezt alkalmazzák a tibetiek), egy másik a vér vörösvértest (és ezen át hemoglobin) koncentrációjának felsrófolása (pl. perui indiánok), egy harmadik pedig a hemoglobin oxigénkötő képességének megvéltoztatása.
Mint azt minden természetjáró tapasztalhatta, a magashegyi környezet alacsony oxigéntartalma elég megterhelő a szervezet számára. Mivel a vér nem tud elegendő oxigént szállítani az izmokhoz, különösen az edzetlen túrázó a legkisebb megterheléstől is lihegni kezd, a legenyhébb szellőtől is elfárad. A probléma kiküszöbölésének egyik útja a rendszeres légzésszám megnövelése (ezt alkalmazzák a tibetiek), egy másik a vér vörösvértest (és ezen át hemoglobin) koncentrációjának felsrófolása (pl. perui indiánok), egy harmadik pedig a hemoglobin oxigénkötő képességének megvéltoztatása.
Utóbbit egy olyan amerikai egérfajban – a szarvasegérben (Peromyscus maniculatus) – figyelték meg, amely populációi a tengerszinttől egészen 4,300 méteres magasságig egyaránt előfordulnak, s így ideális alanyát képezik az összehasonlító populációgenetikai tanulmányoknak.
A szarvasegér két ügyes trükköt "alkalmaz". Egyrészt, más emlősöktől eltérően, a két, egymáshoz közel fekvő α-globin génje (5' és 3') nem teljesen egyforma, hanem ezek egyike ( a 3') egy olyan mutációt hordoz, amely révén megnövekedik az oxigénkötő affinitása. Ennek köszönhetően, az egerek alapból is kétfajta α-globin fehérjével rendelkeznek: egy kisebb affinitásúval, amelyik az oxigéndúsabb, tengerszinthez közelebbi környezetben ideális (mert könnyebben tudja leadni a szövetek oldalán az O2-t), ill. a már említett nagyobb affinitásúval, amely a tegerszinttől távolodva válik egyre fontosabbá. Ez az adaptáció, amelyik mindegyik szarvasegér populációban jelen van, már önmagában is fontos, de egyedül nem tudná megmagyarázni a különböző magasságban élő egerek vérének oxigénmegkötő képessége között észlelhető különbséget.
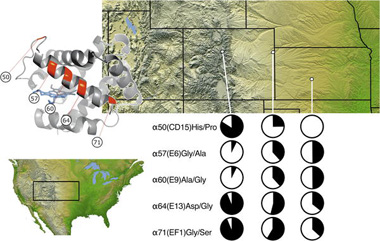 Ehhez szükség van a második "trükkre", amely első pillantásra inkább csak az előző variálásának tűnhet, ugyanis újból olyan mutációkról van szó, amelyek az ominózus oxigénmegkötő képességet befolyásolják. Ezek azonban a másik (5') α-globin gént érinti, azt amelyiknek a mezei egerekben alacsonyabb az oxigenkötő képessége. Ennek a génnek ugyanis több változata (allélja) létezik, attól függően, hogy melyikben hány olyan mutáció fordul elő, amelyik növeli az oxigénmegkötő affinitásukat (és, ne feledjük, ezzel párhuzamosan csökkentik a leadóképességet a szöveteknél). A hegyen élő egerek között igen gyakori, hogy mind az öt, ismert mutáció együtt jelenik meg, míg a tengerszinten élőkben gyakran együtt hiányoznak – a köztes magasságokban pedig átmeneti kombinációk jelennek meg.
Ehhez szükség van a második "trükkre", amely első pillantásra inkább csak az előző variálásának tűnhet, ugyanis újból olyan mutációkról van szó, amelyek az ominózus oxigénmegkötő képességet befolyásolják. Ezek azonban a másik (5') α-globin gént érinti, azt amelyiknek a mezei egerekben alacsonyabb az oxigenkötő képessége. Ennek a génnek ugyanis több változata (allélja) létezik, attól függően, hogy melyikben hány olyan mutáció fordul elő, amelyik növeli az oxigénmegkötő affinitásukat (és, ne feledjük, ezzel párhuzamosan csökkentik a leadóképességet a szöveteknél). A hegyen élő egerek között igen gyakori, hogy mind az öt, ismert mutáció együtt jelenik meg, míg a tengerszinten élőkben gyakran együtt hiányoznak – a köztes magasságokban pedig átmeneti kombinációk jelennek meg.
Mivel az egerek a Sziklás hegység oldalában folyamatosan jelen vannak a Nagy Síkságtól egészen több ezer méterig, elvileg a különböző allélok, az 5' α-globin gén különböző változatai szabadon áramolhatnának a populációk között, hiszen annak semmi akadálya nincs, hogy egy-egy egér élete során pár száz métert kóboroljon valamelyik irányba (és ott szaporodjon). Az allélok jellegzetes rögzülése azonban azt bizonyítja, hogy a természetes szelekció adott magasságban egy adott allélt preferál, s aki mással rendelkezik az hátrányba kerül.
Storz JF, Sabatino SJ, Hoffmann FG, Gering EJ, Moriyama H, et al. (2007) The Molecular Basis of High-Altitude Adaptation in Deer Mice. PLoS Genet 3(3): e45 doi:10.1371/journal.pgen.0030045
 Amikor világot látott szűk másfél éve a kutya genom
Amikor világot látott szűk másfél éve a kutya genom 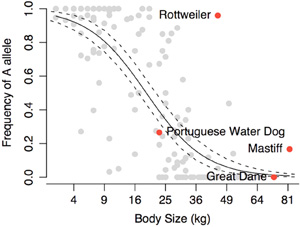 Persze kérdés, hogy mennyiben alkalmazható az egyetlen fajtából szerzett tudás a többiekre. Lehet, hogy a lecsökkent IGF1 szint más, aprónövésű fajtákban is fontos, de az is lehet, hogy azok egyéb okok miatt maradnak kicsik. A kérdést eldöntendő 143 kutyafajtát genotipizált végig a kutatócsoport és az eredmények szerint, a portugál vízikutyában talált „kis” IGF1 allél jelenléte ill. hiánya igen-igen jól korrelál a testmérettel. Jól, de nem tökéletesen (pl. a rottweilerek nagy többségében ez az allél van és mégsem kicsik), éppen ezért helytelen lenne az IGF1-t kinevezni A „méret-génnek”. Mint minden komplex, sok árnyalattal jellemezhető tulajdonság, a testméret is sok géntől függ (azaz poligénes). Az azonban bizonyosnak tűnik, hogy ez közül az egyik kiemelkedően fontos, az IGF1.
Persze kérdés, hogy mennyiben alkalmazható az egyetlen fajtából szerzett tudás a többiekre. Lehet, hogy a lecsökkent IGF1 szint más, aprónövésű fajtákban is fontos, de az is lehet, hogy azok egyéb okok miatt maradnak kicsik. A kérdést eldöntendő 143 kutyafajtát genotipizált végig a kutatócsoport és az eredmények szerint, a portugál vízikutyában talált „kis” IGF1 allél jelenléte ill. hiánya igen-igen jól korrelál a testmérettel. Jól, de nem tökéletesen (pl. a rottweilerek nagy többségében ez az allél van és mégsem kicsik), éppen ezért helytelen lenne az IGF1-t kinevezni A „méret-génnek”. Mint minden komplex, sok árnyalattal jellemezhető tulajdonság, a testméret is sok géntől függ (azaz poligénes). Az azonban bizonyosnak tűnik, hogy ez közül az egyik kiemelkedően fontos, az IGF1. Az élet egy harc, s itt most egészen prózai dologra gondolok: a nap minden órájában szervezetünk (és általában minden állati szervezet) kis betolakodók támadásának van kitéve, legtöbbször anélkül hogy tudnánk róla. Ezek az élősdik rendszertanilag rendkívül különböző csoportokból származnak, egy dologban azonban hasonlítanak egymásra: mind úgy „vélik”, hogy belsőnk ideális körülményeket biztosít életciklusuk valamelyik szakaszához.
Az élet egy harc, s itt most egészen prózai dologra gondolok: a nap minden órájában szervezetünk (és általában minden állati szervezet) kis betolakodók támadásának van kitéve, legtöbbször anélkül hogy tudnánk róla. Ezek az élősdik rendszertanilag rendkívül különböző csoportokból származnak, egy dologban azonban hasonlítanak egymásra: mind úgy „vélik”, hogy belsőnk ideális körülményeket biztosít életciklusuk valamelyik szakaszához. A fejlődés során becsúszó mutációk, az epigenetikai szabályozás és a környezeti hatások miatt, nincs két tökéletesen egyforma élőlény, bár értelemszerűen a klónok, partenogenetikus egyedek és az egypetéjű ikrek meglehetősen közel állnak ehhez. A genetikai anyag közel tökéletes megegyezése az, ami igazán különbözővé teszi pl. utóbbiakat a kétpetéjű ikrektől, amelyek csak éppen annyira hasonlítanak mint egy átlagos testvérpár (hiszen szintén két különböző petesejt és spermium egyesülésével jönnek létre).
A fejlődés során becsúszó mutációk, az epigenetikai szabályozás és a környezeti hatások miatt, nincs két tökéletesen egyforma élőlény, bár értelemszerűen a klónok, partenogenetikus egyedek és az egypetéjű ikrek meglehetősen közel állnak ehhez. A genetikai anyag közel tökéletes megegyezése az, ami igazán különbözővé teszi pl. utóbbiakat a kétpetéjű ikrektől, amelyek csak éppen annyira hasonlítanak mint egy átlagos testvérpár (hiszen szintén két különböző petesejt és spermium egyesülésével jönnek létre).
 A tankönyvi történet szerint, az emlősök mai sokfélesége annak a radiációnak köszönhető, amely mintegy 65 millió évvel ezelőtt, a dinoszauruszok hirtelen kipusztulása után következett be. Egészen addig csak apró éjszakai állatokként húzódtak meg a dinoszauruszok árnyékában, s ha nincs a
A tankönyvi történet szerint, az emlősök mai sokfélesége annak a radiációnak köszönhető, amely mintegy 65 millió évvel ezelőtt, a dinoszauruszok hirtelen kipusztulása után következett be. Egészen addig csak apró éjszakai állatokként húzódtak meg a dinoszauruszok árnyékában, s ha nincs a 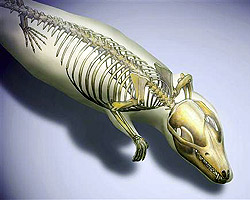 A kínai lelőhelyek továbbra is ontják magukból az érdekesnél érdekesebb fosszíliákat. Most éppen ismét egy ősi emlős került elő, amely egyszerre támasztja alá a középfül evolúciós kialakulásáról szóló elméleteinket és kínál betekintést az evolúció boszorkánykonyhájának egy Hox-gének tüzelte bugyrába.
A kínai lelőhelyek továbbra is ontják magukból az érdekesnél érdekesebb fosszíliákat. Most éppen ismét egy ősi emlős került elő, amely egyszerre támasztja alá a középfül evolúciós kialakulásáról szóló elméleteinket és kínál betekintést az evolúció boszorkánykonyhájának egy Hox-gének tüzelte bugyrába.
 A napfény egyszerre jelent áldást és gondforrást: utóbbit a benne levő
A napfény egyszerre jelent áldást és gondforrást: utóbbit a benne levő  A malária elleni védekezésről beszélve, általában hajlamosak vagyunk egy kalap alá venni magát a malária parazitát és köztes gazdáját, a szúnyogot: a cél mindkettő pusztítása. Persze ez nem meglepő, hiszen a gyakorlatban is elég jól helytáll a "kevesebb szúnyog = kevesebb megbetegedés" egyenlet. De ennek fényében gyakran elfeledkezünk egy apróságról: a Plasmodium a szúnyogoknak is a parazitája, nem csak nekünk, embereknek.
A malária elleni védekezésről beszélve, általában hajlamosak vagyunk egy kalap alá venni magát a malária parazitát és köztes gazdáját, a szúnyogot: a cél mindkettő pusztítása. Persze ez nem meglepő, hiszen a gyakorlatban is elég jól helytáll a "kevesebb szúnyog = kevesebb megbetegedés" egyenlet. De ennek fényében gyakran elfeledkezünk egy apróságról: a Plasmodium a szúnyogoknak is a parazitája, nem csak nekünk, embereknek.
 A
A  Az egerek világa, a szó szoros értelmében, lényegesen színtelenebb, mint a miénk, hiszen az
Az egerek világa, a szó szoros értelmében, lényegesen színtelenebb, mint a miénk, hiszen az {kind=link}
{kind=link}