 Vannak témák, amelyek végigkövetik a blog most már évtizedes történetét, és az egyik legrégebbi történet, aminek szálait göngyölítgetjük, arról szól, hogy az élővilág evolúciója során miként alakult ki az élőlények formagazdagsága.
Vannak témák, amelyek végigkövetik a blog most már évtizedes történetét, és az egyik legrégebbi történet, aminek szálait göngyölítgetjük, arról szól, hogy az élővilág evolúciója során miként alakult ki az élőlények formagazdagsága.
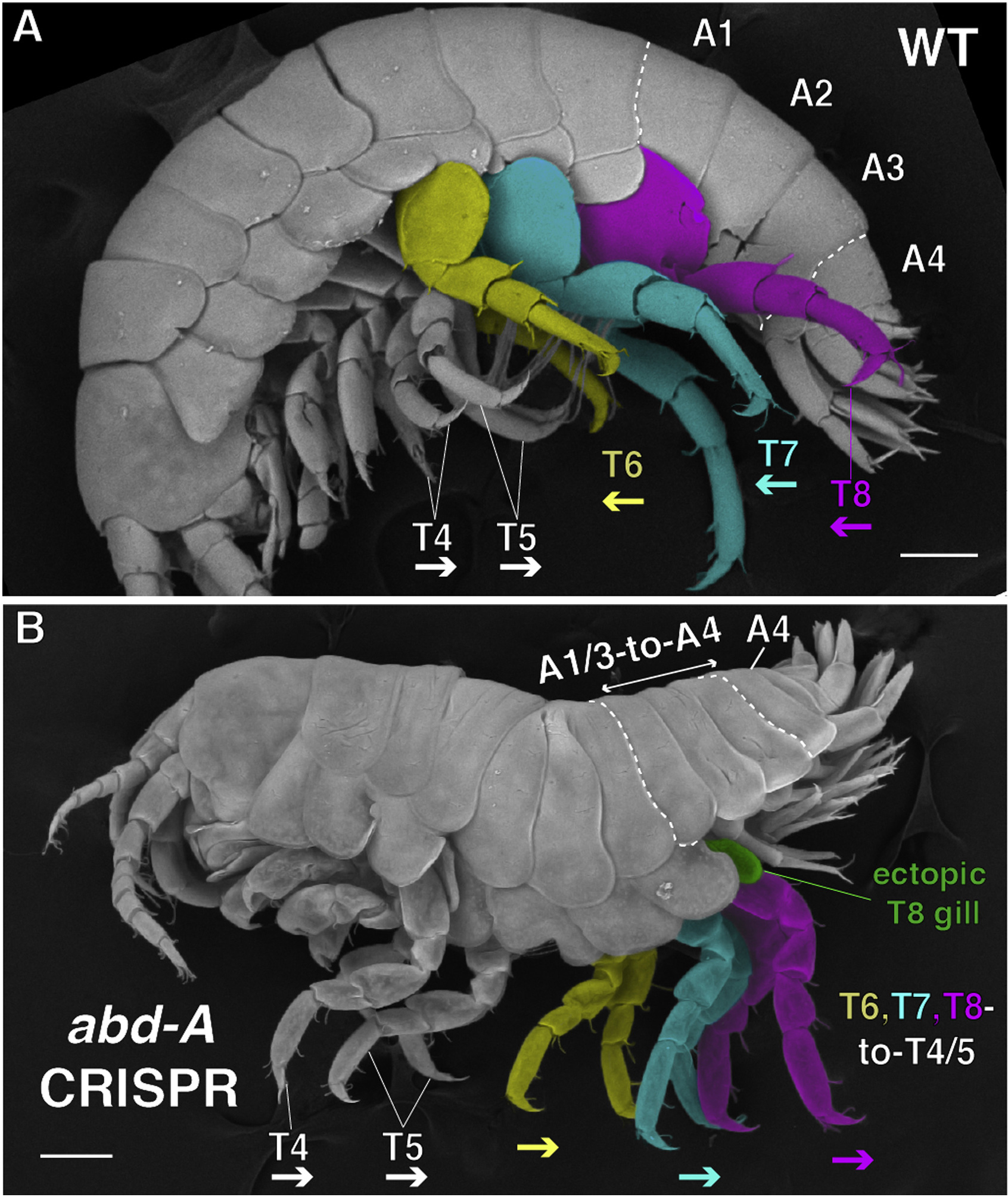
A sztori megkerülhetetlen eleme a testfelépítésért felelős, ún. Hox gének története épp ezért ha van téma, amivel sokszor, sokat foglalkoztunk, ez biztos az. De néha a „sokat” sem elég, főleg, hogy most már jó ideje egyáltalán, szóval időszerű egy kicsit elmerülni ismét abban a megunhatatlan témában, hogy miként formálta és formálja ez az ősi géncsalád például az ízeltlábúak formáit.
Az „alapsztorit” már bő nyolc és fél éve megírtam, és zanzásítva valahogy úgy szól, hogy az ízeltlábúak jól látható szelvényes, ha úgy tetszik „moduláris” felépítését a Hox gének változó expressziós tartománya befolyásolja. Az ízeltlábúak egyes szelvényeinek alapfelépítése tulajdonképpen nagyon egyszerűen megfogalmazható: egy henger alakú testből egy pár nyúlvány lóg ki. Hogy ez a nyúlvány járólábként, szájszervként, vagy csápként, az a szelvény pozíciójától függ. És ezt a pozicionális információt a Hox gének határozzák meg, amelyek ha elromlanak, a szelvény identitása is elromlik, így szájszervek, vagy csápok helyett péládul járóábak jelennek meg a fej szelvényein is (ezeket nevezzük homeotikus mutációknak).
Amikor aztán sokan azt is megfigyelték, hogy az egyes fontosabb Hox gének kifejeződése nagyobb taxonómiai csoportonként jellegzetes változásokat hordoz, sokan elkezdték fejtegetni, hogy valószínűnek tűnik, sok esetben a csoportok sztereotipikus külalakjában megfigyelhető jellegezetes különbségek is az ilyen Hox-expresszióban jelentkező különbségekre vezethetők vissza. Magyarán az evolúció szívesen „játszott” ezekkel a génekkel.
Egy kattintás ide a folytatáshoz…. →

 Bár laikusok számára a halak kifejezetten unalmas jószágoknak is tűnhetnek, egy biológus számára megunhatatlan tárházát nyújtják az érdekességeknek. Mégpedig azért, mert a gerinces fajok kb. fele hal, ennek megfelelően elképesztő forma-, életmód- és viselkedésgazdagságot rejt ez a csoport.
Bár laikusok számára a halak kifejezetten unalmas jószágoknak is tűnhetnek, egy biológus számára megunhatatlan tárházát nyújtják az érdekességeknek. Mégpedig azért, mert a gerinces fajok kb. fele hal, ennek megfelelően elképesztő forma-, életmód- és viselkedésgazdagságot rejt ez a csoport.
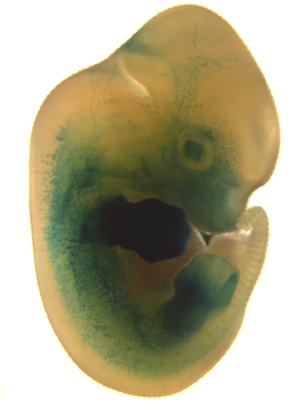 Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.
Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.

 Jó pár évvel ezelőtt, a blog hajnalán, az egyik legizgalmasabb és legfelkapottabb kérdés a biológiában az volt, hogy a különböző fajok (alfajok) morfológiai sajátosságai miképpen alakulnak ki a fejlődés során. Az evodevo tudománya – mert arról van szó – azóta kicsit kiment a mainstream divatból, de ha engem kérdezne bárki, szerintem továbbra is az egyik legizgalmasabb kérdés, amivel biológus foglalkozhat.
Jó pár évvel ezelőtt, a blog hajnalán, az egyik legizgalmasabb és legfelkapottabb kérdés a biológiában az volt, hogy a különböző fajok (alfajok) morfológiai sajátosságai miképpen alakulnak ki a fejlődés során. Az evodevo tudománya – mert arról van szó – azóta kicsit kiment a mainstream divatból, de ha engem kérdezne bárki, szerintem továbbra is az egyik legizgalmasabb kérdés, amivel biológus foglalkozhat. A barlangi élet különös alkalmazkodásokat követel meg azoktól az élőlényektől, akik a létnek ezt a sötét formáját választják. A jóformán örök sötétségben fölöslegessé válik a felszínen oly fontos látás, lényegtelenné a bármilyenfajta mintázat, más érzékek és más jellegek válnak fontossá az életbenmaradás és fajfenntartás céljából.
A barlangi élet különös alkalmazkodásokat követel meg azoktól az élőlényektől, akik a létnek ezt a sötét formáját választják. A jóformán örök sötétségben fölöslegessé válik a felszínen oly fontos látás, lényegtelenné a bármilyenfajta mintázat, más érzékek és más jellegek válnak fontossá az életbenmaradás és fajfenntartás céljából. 