Hogy már rögtön az egész blog legelején lelőjem a legnagyobb evolúciós közhelyet: Theodosius Dobzhansky, minden idők egyik legnagyobb evolúcióbiológusának szavaival élve: ”A biológiában semminek nincs értelme, kivéve ha az evolúció fényében nézzük.” Közhely, de nagyon igaz, főleg napjainkban, amikor a fejlődésbiológia, evolúcióbiológia és genetika összefonódásából egy új tudományág, az ún. EvoDevo van születőben.
A darwini alapgondolat a közös őstől való leszármazás, azaz minden ma létező élőlény eredete visszavezethető egyetlen régen élt ősi organizmushoz (ez az, amit a szakirodalom Universal Common Ancestor – UCA néven ismer). Ebből az ősi, primitív szervezetből jött létre a ma ismert élővilág sokfélesége a természetes szelekció viszonylag egyszerű logikája alapján. Ezen elegáns gondolatmenet egyik alapkövét Karl Ernst von Baer, német embriológus törvényei képezték, amelyek (durva közelítésben) azt fogalmazták meg, hogy az egyedfejlődés során az általános jegyek hamarabb, míg a specifikus jellegzetességek később jelennek meg (azaz minél közelebb álló fajokat vizsgálunk, annál tovább hasonló az embrionális fejlődésük). Továbbgondolva, ebből az is világosan következik, hogy a különböző fajok közötti különbségek legelőször az egyedfejlődés valamelyik szakaszában jelennek meg. Vagyis ha meg akarjuk érteni az evolúciót akkor érdemes beleásnunk magunkat a fejlődésbiológiába (és természetesen vice versa). Sean B. Carroll, az evodevo-s társaság egyik vezéralakja legutóbbi könyvének (”Endless Forms Most Beautiful”) előszavában megpróbálja zanzásítani az új terület esszenciáját, több-kevesebb sikerrel: „A formák közötti különbségek az egyes gének felhasználási helyét és idejét befolyásoló evolúciós változásokból erednek, különös tekintettel azokra a génekre, amelyek bizonyos struktúrák számát, alakját vagy méretét befolyásolják.” Nnna, egész tömören tehát valami ilyesminek a vizsgálatával foglalkozik az EvoDevo. Hogy a meghatározás mögött mi rejlik, azt majd a blog keretén belül igyekszem/-szünk górcső alá venni.
Az evolúciós elmélet fontos következménye, hogy a természet hozott anyagból dolgozik, azaz már létező élőlények adaptálódnak új körülményekhez. Ez számos esetben azzal jár, hogy olyan szervek, amelyek valamelyik ős számára létfontosságúak voltak, hirtelen feleslegessé válnak, s ezért lassan elsatnyulnak, elcsökevényesednek. Az darwini gondolat ezen ütős bizonyítékait nevezzük csökevény (vagy vesztigiális) szerveknek, s ez a kis sorozat néhány jellegzetesebb (és érdekesebb) bemutatásával fogja az időt pocsékolni.
Mint a címben diszkréten próbáltam rá utalni, ebben az első részben a kígyók (hiányzó) lábáról lesz szó, hogy hova tűnt el és miért.
 1. Ábra: A mellső- és hátsó végtagok kialakulását befolyásoló transzkripciós faktorok (forrás: [2]).
1. Ábra: A mellső- és hátsó végtagok kialakulását befolyásoló transzkripciós faktorok (forrás: [2]).
Ha a kígyók rendszertani besorolását nézzük, akkor elég egyértelmű, hogy hüllőként a gerincesek Tetrapoda (azaz „Négylábúak”) csoportjához tartoznak. Azonban az is egyértelmű, hogy nincs négy lábuk. A kérdés adott: miért?, ám hogy megértsük, ahhoz először egy végtagkialakulás-gyorstalpalón kell átesnünk. Az egyes végtagok pozícióját a gerincesek testének hosszanti felosztásában kulcsszerepet játszó homeobox (hox) gének kombinációja határozza meg (kisebb betű+szám kombináció előrébb található, míg nagyobb betű+szám hátsóbb expressziós területet jelöl). Például a mellső végtagok a hoxc6 gén expressziós területének kezdete előtt találhatók (lásd 2. ábra). A végtagkialakulás pozíciójában a hámréteg (epidermisz) alatt mezodermális sejtek gyűlnek össze és az ezek által termelt növekedési faktor (FGF10) váltja ki az epidermisz és az alatta található szövetek közötti interakciót. Ennek következményeként specifikus transzkripciós faktorok lépnek működésbe az első és a hátsó végtag leendő pozíciójában (Tbx5 a mellső végtagok esetén, illetve Tbx4 és Pitx1 a hátsó végtagokban (lásd 1. ábra) [1,2]) melyek elindítják a „végtagbimbók” (limb buds) növekedését. Ezekben a végtag-kezdeményekben további bonyolult felosztási folyamatok játszódnak le (ezekről majd valamikor később – talán – lesz szó bővebben), de a legfontosabb, hogy hátsó részükön kialakul az úgynevezett polarizáló aktivitás zónája (zone of polarizing activity – ZPA), amely koordinálni fogja ezt a felosztást az általa termelt sonic-hedgehog (shh) nevű molekula révén.
Na, akkor ennyi fejtágítás után lássuk, hogy mi is történik (pontosabban nem történik) a kígyók embrionális fejlődése során.
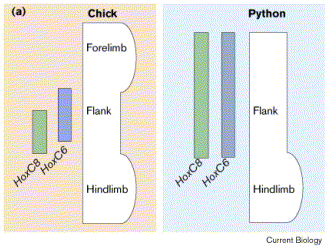 2. Ábra: A kígyókban egyes hox gének expressziós tartománya megváltozott. (forrás: [3])
2. Ábra: A kígyókban egyes hox gének expressziós tartománya megváltozott. (forrás: [3])
Az első (témánk számára) fontos változás, hogy néhány karakterisztikus hox gén expressziója megváltozott: a hoxb5, hoxc6 és hoxc8 szinte a test teljes hosszában kifejeződnek, különösen feltűnő, hogy expressziós tartományuk elülső határa a fejnél található (lásd 2. ábra). Ez kettős következménnyel jár: egyrészt megszűnik az elülső végtag kialakulásához szükséges pozicionális információ (mert nincs többé olyan, hogy „hoxc6 előtt” a gerincoszlop mentén), másrészt, mivel ezen hox gének kombinációja (más gerincesekben is) háti identitást eredményez, nyilvánvaló, hogy a nyaki csigolyák hátivá váltak, valamint a kígyók megnyúlt teste leginkább egy hosszú hátnak tekinthető [4]. Mellső végtag (hiánya): kipipálva.
A hátsó végtag egy kicsit bonyolultabb történet. Bonyolultabb, mert e tekintetben kígyó és kígyó között is akad különbség. Míg a rendszertani besorolásukat tekintve a Boidae alcsaládba tartozó boák és pythonok esetében felfedezhetünk hátsó végtag maradványokat, addig más kígyók esetében már ilyesmivel sem találkozunk.
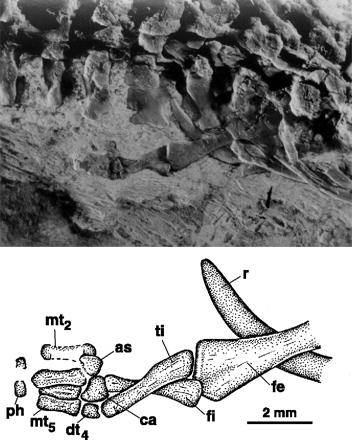 3.Ábra: A 95 millió éve élt Haasiophis terrasanctusnak jól felismerhető hátsó lába volt. (forrás: [5])
3.Ábra: A 95 millió éve élt Haasiophis terrasanctusnak jól felismerhető hátsó lába volt. (forrás: [5])
Ez minden valószínűség szerint a Boidae alcsalád ősiségét bizonyítja, hiszen a fosszilis anyagban fellelhetünk olyan kígyócsontvázakat, amelyeknek bár csökevényes, de jól felismerhető hátsó- (és csak hátsó!) végtagja van. Ilyen például a kb. 95 millió éves Haasiophis terrasanctus (3. ábra) amelyet sokan egy átmeneti láncszemnek tekintenek a kígyók négylábú, gyíkszerű őse és a mai kígyók között [5]. (Ugyanakkor itt illendő hangsúlyozni, hogy tudásunk még elég hiányos kígyó fosszíliák terén, így nem egyértelmű, hogy a Haasiophis valóban a boák ősének tekinthető-e, sőt az sem kizárt, hogy a hátsó láb elvesztése többször, egymástól függetlenül következett be az idők során a kígyók családjában.) A boák embrionális fejlődése során a hátsóvégtag-„bimbó” ugyan megjelenik a testfalon, azonban valamiért mégsem hoz létre teljes értékű végtagot. Hogy miért nem arra a génexpressziós vizsgálatok adtak magyarázatot: valamiért (és ez még pontosan nem tisztázott) a shh gén nem kapcsolódik be az alakuló hátsó végtag ZPA területén. Ennek következtében természetesen az általa irányított folyamatok sem kezdődnek el, ergo a boáknak nem lesz rendes lába. S ami még érdekesebbé teszi ezt, az az, hogy a jelek szerint a végtag-”bimbó” megfelelő jelek esetén képes lenne létrehozni egy végtagot, mert amennyiben csirkeembrióba átültetik a kígyó végtagkezdeményét az működőképes lesz és szerepet játszik a végtag kialakításában. Ezzel tömören le is rendeztük a boák hátsó lábát, de mi a helyzet a többi kígyó esetén? A válasz kicsit kiábrándító: még nem tudjuk (az evodevo sajnos nem az a terület, ahova sok K+F pénz folyik, így viszonylag kevés kísérletre van lehetőség, azok viszont nagyot szólhatnak ;-)). Csak találgatni tudunk, de mivel egyáltalán nincs hátsóvégtag-kezdemény, sokan azt valószínűsítik, hogy szintén a Hox-kód változása lehet a dologban. Remélhetőleg valaki előbb-utóbb a dolog végére jár, akárcsak annak, hogy hasonló változások vezettek a lábatlan gyíkok végtagjainak eltűnéséhez, vagy netán a végtagkialakulási folyamat más pontjain következtek be változások.
[1] Minguillon, C., Del Buono, J. and Logan, M.P. (2005) Tbx5 and Tbx4 are not sufficient to determine limb-specific morphologies but have common roles in initiating limb outgrowth. Developmental Cell 8(1):75-84.
[2] Graham, A. and McGonnell. I. (1999) Limb development: Farewell to arms. Current Biology 9(10):R368-370.
[3] Graham, A. and McGonnell. I. (1999) Developmental evolution: This side of paradise. Current Biology 9(17):R630-632.
[4] Cohn, M. J. and Tickle, C. (1999) Developmental basis of limblessness and axial regionalization in snakes. Nature 399: 474-479.
[5] Tchernov, E., Rieppel, O., Zaher, H., Polcyn, M.J. and Jacobs, L.L. (2000) A fossil snake with limbs. Science 287(5460): 2010-2012.