
 A szexuális szaporodás elengedhetetlen feltétele, hogy két, ellenkező nemű gamétát hordozó egyed valamiféleképpen összehozza ezeket az ivarsejteket. Ehhez nem árt valamiféleképpen szinkronizálni a gaméták kibocsájtását, különösen, ha egyesülésüket nem segítjük elő a (mint a mi esetünkben) belső megtermékenyítés biztosította fizikai közelséggel.
A szexuális szaporodás elengedhetetlen feltétele, hogy két, ellenkező nemű gamétát hordozó egyed valamiféleképpen összehozza ezeket az ivarsejteket. Ehhez nem árt valamiféleképpen szinkronizálni a gaméták kibocsájtását, különösen, ha egyesülésüket nem segítjük elő a (mint a mi esetünkben) belső megtermékenyítés biztosította fizikai közelséggel.
Egyes csalánozó fajok, mint a képen látható Cladonema, ezt úgy oldották meg, hogy egyszerre bocsájtják ki ivarsejtjeiket, hajnalban ill. alkonyatkor. Az adott napszakhoz köthető ívás időzítését elvileg kétéleképpen lehet megoldani: vagy egy belső óra segítségével mérjük a hátralevő időt, vagy pedig érzékeljük az adott napszakra jellemző fényviszonyokat és ennek függvényében indul a "tánc".
A Cladonema feltehetőleg az utóbbi megoldást választotta és ezt azért gondoljuk így, mert az ivarszervei környékén különleges, opszin tartalmú sejtek találhatók. Az opszinok fényérzékeny molekulák, amelyek kialakulása minden valószínűség szerint szorosan köthető a többsejtű állatok "fényérzékelő szervének" (azaz szemének) megjelenéséhez. Ennek megfelelően elsődleges előfordulási helyük a különböző szemek fényérzékeny sejtjeiben (fotoreceptoraiban) van. S ez épp annyira igaz a csalánozókra (akiknek meglepően komplex szemük van), mint ránk, emberekre.
Ezért is különleges a Cladonema ivarszerv-specifikus opszinja, amely a szekvenciavizsgálatok szerint a csalánozó szem-specifikus opszinjaiból jött létre az evolúció során. Egyébként ez a medúza igen csak bőven ellátott opszinokból: Walter Gehring csoportja nem kevesebbet, mint tizenyolcat lelt benne, a legtöbb szerepéről . Vizsgálatuk egyetlen (nagy) hiányossága, hogy az opszin tartalmú sejtek ívásban betöltött szerepét ők is leginkább csak sejtik a pozíciójuk alapján, az igazi funkcionális vizsgálatok (még ?) hiányoznak. A minimum az lett volna, hogy az új opszinok spektrális tulajdonságait górcső alá veszik, hogy valóban a hajnali/esti fény aktiválja-e ezeket a fehérjéket, de sajnos erre nem került sor. Szóval reméljük, még lesz folytatás, mert a téma érdekes.
Suga H, Schmid V, Gehring WJ (2008) Evolution and functional diversity of jellyfish opsins. Curr Biol 18(1): 51-55.
 A második világháború után, a japán frontról visszatérő
A második világháború után, a japán frontról visszatérő 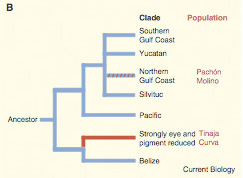 Az elvi részen túl persze még azt sem tudjuk pontosan, hogy az egyes halakban milyen mutáció okozza a szem elsorvadását, mindenesetre mivel számos vak Astyanax populáció nagyon távol él egymsától, feltételezhető, hogy a párhuzamos konvergens evolúció klasszikus példájával állunk szemben, vagyis a vakság egymástól függetlenül alakult ki. Ennek tesztelésére tökéletesen alkalmas a komplementáció, épp ezért végezte ezt el az NYU egyik kutatója, Richard Borowsky.
Az elvi részen túl persze még azt sem tudjuk pontosan, hogy az egyes halakban milyen mutáció okozza a szem elsorvadását, mindenesetre mivel számos vak Astyanax populáció nagyon távol él egymsától, feltételezhető, hogy a párhuzamos konvergens evolúció klasszikus példájával állunk szemben, vagyis a vakság egymástól függetlenül alakult ki. Ennek tesztelésére tökéletesen alkalmas a komplementáció, épp ezért végezte ezt el az NYU egyik kutatója, Richard Borowsky. Bár a növények (részben hozzánemértésünk miatt) szembeötlően kevés figyelmet kaptak eddig a blogban, most részleges kivételt tennék, pont egy egyébként eléggé indulat-generáló, bioetikai kérdés kapcsán.
Bár a növények (részben hozzánemértésünk miatt) szembeötlően kevés figyelmet kaptak eddig a blogban, most részleges kivételt tennék, pont egy egyébként eléggé indulat-generáló, bioetikai kérdés kapcsán.
 A napokban új, ingyenesen elérhető havilap indult az evolúció iránt érdeklődő laikusok, és a tárgyat oktató tanárok számára,
A napokban új, ingyenesen elérhető havilap indult az evolúció iránt érdeklődő laikusok, és a tárgyat oktató tanárok számára,  Indiánokról írni általában véve hálás feladat, hiszen a téma természeténél fogva, mindig akad egy kis egzotikum, vadregényes tájak, számunkra furcsa kinézetű és viselkedésű emberek. És mint nagy írónk ;-)) May Károly esete is bizonyítja, még csak el sem kell utazni az amerikai kontinensre, hogy a témáról bestsellereket írjunk.
Indiánokról írni általában véve hálás feladat, hiszen a téma természeténél fogva, mindig akad egy kis egzotikum, vadregényes tájak, számunkra furcsa kinézetű és viselkedésű emberek. És mint nagy írónk ;-)) May Károly esete is bizonyítja, még csak el sem kell utazni az amerikai kontinensre, hogy a témáról bestsellereket írjunk.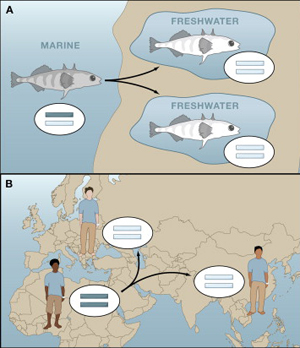 Az elmúlt év legvisszatérőbb témája az állati szőrszínezet, vagyis pigmentáció evolúciója volt, így talán stílszerű, hogy az év utolsó postjának tervezett, de persze az új évbe átcsúszó beírás is ezzel a témával foglalkozzon.
Az elmúlt év legvisszatérőbb témája az állati szőrszínezet, vagyis pigmentáció evolúciója volt, így talán stílszerű, hogy az év utolsó postjának tervezett, de persze az új évbe átcsúszó beírás is ezzel a témával foglalkozzon. Elsőre elbűvölően egyszerűnek, logikusnak és megnyerőnek hangzik az okfejtés: ahogy az ember egyre inkább megtanulta leigázni a természetet és eszközei révén alakítani azt, úgy vonta ki magát is a természeti törvények legalapvetőbbike, a természetes szelekció alól. Ha viszont ez így van, annak fel kellene leljük a nyomát a genomunkban is, hiszen ha az elmúlt kb. 50.000 év során már nem a szelekció formálta génjeinket, akkor az említett intervallumban lényegesen kevesebb adaptív (hasznos) változás rögzült az emberiség genetikai állományokban, mint az azt megelőző időszakban.
Elsőre elbűvölően egyszerűnek, logikusnak és megnyerőnek hangzik az okfejtés: ahogy az ember egyre inkább megtanulta leigázni a természetet és eszközei révén alakítani azt, úgy vonta ki magát is a természeti törvények legalapvetőbbike, a természetes szelekció alól. Ha viszont ez így van, annak fel kellene leljük a nyomát a genomunkban is, hiszen ha az elmúlt kb. 50.000 év során már nem a szelekció formálta génjeinket, akkor az említett intervallumban lényegesen kevesebb adaptív (hasznos) változás rögzült az emberiség genetikai állományokban, mint az azt megelőző időszakban.
 A cetek Darwin óta mindig is különleges kis helyet foglaltak el az evolúcióval foglalkozó kutatók szívében, hiszen ez az az emlős csoport, amely a legügyesebbnek bizonyult a vízi környezet meghódításában – olyannyira, hogy mára már képtelenek rövidebb időt is a parton tölteni.
A cetek Darwin óta mindig is különleges kis helyet foglaltak el az evolúcióval foglalkozó kutatók szívében, hiszen ez az az emlős csoport, amely a legügyesebbnek bizonyult a vízi környezet meghódításában – olyannyira, hogy mára már képtelenek rövidebb időt is a parton tölteni.
 18O izotóp tartalma vízi életmódot sejtet és ezt támasztja alá megvastagodott külső rétegük is, amely ballasztként funkcionált (hasonlóan számos jelenkori, vízi életmódot folytató emlőshöz). A lábak ugyanakkor nem úszáshoz szoktak, így Thewissenék azt feltételezik, hogy egy ősi növényevőről van szó, amelyet a ragadozóktól való félelem készetett a vizes környezethez való átszokásra, de enni még ki-kijárt a szárazföldre – akárcsak a vízilovak. S ha ez így volt, a következő fontos – talán A legfontosabb – evolúciós lépés lehetett a a cetek története során az étrend átalakulása, a húsevésre való áttérés. De ez már a
18O izotóp tartalma vízi életmódot sejtet és ezt támasztja alá megvastagodott külső rétegük is, amely ballasztként funkcionált (hasonlóan számos jelenkori, vízi életmódot folytató emlőshöz). A lábak ugyanakkor nem úszáshoz szoktak, így Thewissenék azt feltételezik, hogy egy ősi növényevőről van szó, amelyet a ragadozóktól való félelem készetett a vizes környezethez való átszokásra, de enni még ki-kijárt a szárazföldre – akárcsak a vízilovak. S ha ez így volt, a következő fontos – talán A legfontosabb – evolúciós lépés lehetett a a cetek története során az étrend átalakulása, a húsevésre való áttérés. De ez már a 
 Hát akkor Foxp2, vagyis a „beszéd-gén”. Már megint, hiszen nemrég, a gén neandervölgyi megfelelőjének kapcsán
Hát akkor Foxp2, vagyis a „beszéd-gén”. Már megint, hiszen nemrég, a gén neandervölgyi megfelelőjének kapcsán  Az emberi beszéd, pontosabban annak a tanult hangképzéshez kapcsolódó elemének tanulmányozására, a laikusok talán váratlan módon egyáltalán nem főemlősöket használnak a kutatók. Sőt mégcsak nem is emlősöket, hanem énekesmadarakat, azok közül is az állatkereskedések kedvelt zebrapintyét. Teszik ezt azért, mert a madarak éneke jellegében emlékeztet a mi kommunikációnkra: rövidebb részekből, „szavakból” felépülő „mondatok” különíthetők el benne. A „szavakat” a fiatalabb madarak az idősek „mondatainak” utánzásával sajátítják el, majd felnőtt korukban maguk is képesek lesznek valamennyire variálni őket.
Az emberi beszéd, pontosabban annak a tanult hangképzéshez kapcsolódó elemének tanulmányozására, a laikusok talán váratlan módon egyáltalán nem főemlősöket használnak a kutatók. Sőt mégcsak nem is emlősöket, hanem énekesmadarakat, azok közül is az állatkereskedések kedvelt zebrapintyét. Teszik ezt azért, mert a madarak éneke jellegében emlékeztet a mi kommunikációnkra: rövidebb részekből, „szavakból” felépülő „mondatok” különíthetők el benne. A „szavakat” a fiatalabb madarak az idősek „mondatainak” utánzásával sajátítják el, majd felnőtt korukban maguk is képesek lesznek valamennyire variálni őket.