
 “A kevesebb több lett volna”, írta Szendi Gábor kritikámra adott válaszáról egyik pártolója, de szerencsére nem így lett. Ha ugyanis kevesebb lett volna, akkor kevésbé tudnám Szendi ezen írását a paleolit-diétát népszerűsítő tevékenysége állatorvosi lovaként használni, így viszont tökéletesen visszaigazolja előzőleg megfogalmazott kritikáimat: 1.) “a Nyugati táplálkozás tudományos kritikája” nem más mint egy nagyon szelektív (és emiatt torzító) merítés a rendelkezésünkre álló szakmai irodalomból, ill. 2.) Szendi genetikai ismeretei igencsak felszínesek.
“A kevesebb több lett volna”, írta Szendi Gábor kritikámra adott válaszáról egyik pártolója, de szerencsére nem így lett. Ha ugyanis kevesebb lett volna, akkor kevésbé tudnám Szendi ezen írását a paleolit-diétát népszerűsítő tevékenysége állatorvosi lovaként használni, így viszont tökéletesen visszaigazolja előzőleg megfogalmazott kritikáimat: 1.) “a Nyugati táplálkozás tudományos kritikája” nem más mint egy nagyon szelektív (és emiatt torzító) merítés a rendelkezésünkre álló szakmai irodalomból, ill. 2.) Szendi genetikai ismeretei igencsak felszínesek.
Mielőtt azonban folytatnám Szendi gondolatmenetének a kritikáját, egy, a kommentekben többször előbukkanó félreértést szeretnék eloszlatni. Többen úgy értelmezték felvetéseimet, hogy azokban kétségbe vonom a paleolit-diéta működőképességét. Ezt azonban sehol nem írtam. Valóban csak olyan cikkekről írtam, amelyek közvetve, vagy közvetlenül cáfolják a paleolit diéta fontos téziseit, de elsősorban azért mert ezeket Szendi elmulasztotta ismertetni, én pedig szeretném, hogy mindenki lássa, van az éremnek egy másik oldala is: a dietetikában kevés valóban fekete és fehér dolog van, általában a szürke különböző árnyalataival találkozunk. Nem bagatellizálom a személyes sikertörténeteket, amelyekből tucatnyi előkerült: készséggel hiszem el, hogy sok kommentelőnek konkrétan a mértékletes táplálkozás ezen formája vált be. Azt vonom csak kétségbe, hogy az anekdotikus esetek akár együttesen is valamiféle statisztikailag jelentős bizonyítékot jelentenének arra nézve, hogy a visszafogott étkezés más formái ne működ(het)nének, vagy hogy a “Paleolit táplálkozásban” leírt gabona- és tej-ellenes kirohanások, abban a formában igazak lennének. Nyilván sokkal jobban el lehet adni egy ilyen témájú könyvet, hogy ha azt írjuk rá, hogy “ez az egy igaz járható út, kövesd, vagy korán meghalsz”, mint a visszafogottabb “ez egy x+1. diéta recept, ha más nem vált be önnek, próbálja ki és hátha” szöveggel. Viszont ettől még az előbbi nem lesz igaz.
 Új hónap, új meetup. Ezúttal október 20-án, a
Új hónap, új meetup. Ezúttal október 20-án, a 
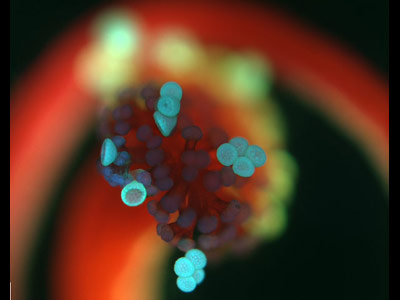
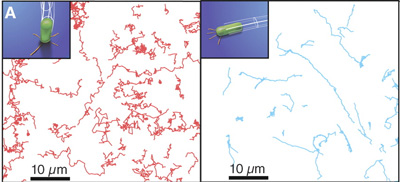 Bakteriális mozgás kapcsán leggyakrabban azokra az ostor által katalizált helyváltaztotásokra gondolunk, amelyeknek SexComb kolléga olyan
Bakteriális mozgás kapcsán leggyakrabban azokra az ostor által katalizált helyváltaztotásokra gondolunk, amelyeknek SexComb kolléga olyan  pilA mutánsok esetében nem jelentkezett, s ennek eredményeként ott a normális esetben megfigyelhető egyenletes biofilmek helyett, csomók keletkeztek.
pilA mutánsok esetében nem jelentkezett, s ennek eredményeként ott a normális esetben megfigyelhető egyenletes biofilmek helyett, csomók keletkeztek.
 A mexikói vaklazac (Astyanax mexicanus) mindig hálás téma, ha evo-devo témakörben írogatunk, most pedig a szokásost még egy kis viselkedés-biológiával is kiszínezhetjük.
A mexikói vaklazac (Astyanax mexicanus) mindig hálás téma, ha evo-devo témakörben írogatunk, most pedig a szokásost még egy kis viselkedés-biológiával is kiszínezhetjük.
