
Nem akarom elvenni dolphin kenyerét, de nem tudtam megállni, hogy bele ne kontárkodjak a HOX génekbe, hiszen nemrég egy roppant érdekes közlemény jelent meg.
Azt ugye tudjuk, hogy a HOX gének alakítják ki a testszelvények különbségeit, ezen gének termékei transzkripciós faktorok, amelyek sok más gén kifejeződését szabályozzák. Az is tiszta sor, hogy az új testfelépítésű élőlények kialakulása tulajdonképpen a HOX gének drámai változásaival járt együtt. Az viszont eddig kérdéses volt, hogy ha a HOX gének ilyen alapvető fontosságú feladatot töltenek be, hogyan változhatnak mégis? Ahogy a közlemény szerzői megfogalmazták a két kérdésüket:
– Hogyan változnak a szabályozógének az evolúció során, hogy különböző élőlények egyedfejlődését irányítsák?
– Hogyan bírják ki az élőlények ezeket a változásokat, ha vélhetőleg igen súlyos következményekkel járnak?
Nem akarom elvenni dolphin kenyerét, de nem tudtam megállni, hogy bele ne kontárkodjak a HOX génekbe, hiszen nemrég egy roppant érdekes közlemény jelent meg.
Azt ugye tudjuk, hogy a HOX gének alakítják ki a testszelvények különbségeit, ezen gének termékei transzkripciós faktorok, amelyek sok más gén kifejeződését szabályozzák. Az is tiszta sor, hogy az új testfelépítésű élőlények kialakulása tulajdonképpen a HOX gének drámai változásaival járt együtt. Az viszont eddig kérdéses volt, hogy ha a HOX gének ilyen alapvető fontosságú feladatot töltenek be, hogyan változhatnak mégis? Ahogy a közlemény szerzői megfogalmazták a két kérdésüket:
– Hogyan változnak a szabályozógének az evolúció során, hogy különböző élőlények egyedfejlődését irányítsák?
– Hogyan bírják ki az élőlények ezeket a változásokat, ha vélhetőleg igen súlyos következményekkel járnak?
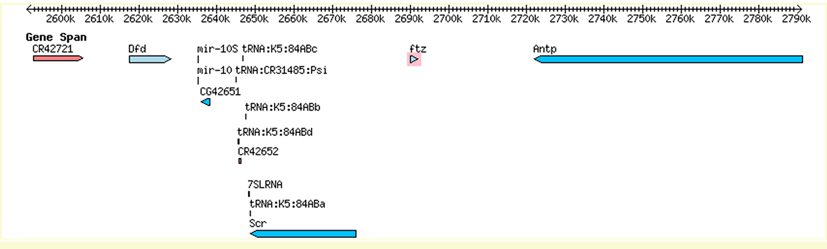
A vizsgálati rendszerük a Fushi tarazu (Ftz) fehérje, amely az aminosavsorrendje alapján egy homeotikus fehérje, az Antennapedia (Antp) fehérjééhez nagyon hasonló homeoboxot tartalmaz, ám meglepő módon nem HOX fehérjeként működik, hanem pair-rule génként. Ez azért is meglepő, mivel in vitro, az izolált Ftz fehérje képes DNS –t kötni, pont úgy, ahogy az Antp. Ráadásul a genomban közvetlenül az Antp és az Scr (Sex combs reduced, egy másik HOX gén) között található a ftz gén. Az ftz gén az embrionális fejlődés nagyon korai szakaszában fejeződik ki, hét, a testtengelyre merőleges sávban, hiányában az egyed életképtelen, tehát mindenképpen valamilyen fontos szerepet tölt be. Nagyon úgy néz ki, hogy az Ftz fehérje jó példa a molekuláris evolúció vizsgálatára. Hogyan jöhetett létre ez a furcsa fehérje?
Az új gének kialakulásának szokásos modellje, hogy egy duplikáció nyomán egy gén megkettőződik, majd az egyik példány mutációkat gyűjt be, így megváltozik. Ebben az esetben úgy tűnik, pontosan ez a helyzet, az Ftz fehérje homodoménje mindössze két aminosavban tér el az Antp fehérje homeodoménjétől, míg az Scr fehérjéé négy ponton különbözik, ráadásul a közvetlen közelükben található az őt kódoló gén is, összességében véve az Ftz fehérje nagyon úgy néz ki, mint egy módosult Antp. Ahhoz hogy ez a változás lehetővé váljon, szükséges volt, hogy gyengüljön a génre nehezedő szelekciós nyomás. Vélhetőleg az eredeti duplikációval keletkezett az ftz és az Antp gén, az általuk kódolt két fehérje részben vagy egészben átfedő működést végzett, így az ftz gén megváltozhatott, hiszen az Antp gén terméke is elegendő volt az életben maradáshoz.
Ezt mutatja, hogy számos más rovar Fushi tarazu fehérjéje az Antennapedia fehérjéhez hasonlóan működik, ha Drosophila melanogasterbe juttatták őket. Például, ha a Drosophila melanogaster lárvában túltermeltetik az Antp fehérjét, a kikelő legyek csáp helyett lábakat viselnek majd, ez egy közismert fenotípus. Ha ugyanígy túltermeltetik az ecetmuslica Ftz fehérjéjét, a csápok csápok maradnak. Azonban, ha a Tribolium castaneum nevű bogár Ftz fehérjéjét termeltették túl ugyanígy, a kikelő legyek csápjai helyett lábak nőttek, bár ez a fenotípus gyengébb volt, mint az Antp túltermelésével kapott, de ez az Ftz ortológ még képes valamilyen mértékig helyettesíteni az Antp fehérjét.
Az ismert, hogy ha egy ecetmuslica embrióban túltermeltetik az Ftz fehérjét, a testszelvények közül hiányozni fognak azok a szelvények, amelyek a vad típusú állatban nem fejezik ki az Ftz fehérjét. Az Antp és az Scr túltermeltetése nyomán semmilyen hasonló hatást sem tapasztaltak. Viszont ha túltermeltetik a T. castaneum Ftz ortológját, hasonló fenotípust mutat, mint a D. melanogaster Ftz fehérjéje, csak gyengébbet. Ebből a kísérletből az látszik, hogy a Tc-Ftz fehérje nem egyszerűen egy homeotikus fehérje, valamilyen mértékig képes helyettesíteni a Dm-Ftz fehérjét is, teljesen olyan, mint egy átmeneti alak a tisztán homeotikus működésű ősi Ftz és a tisztán szegmentációs működésű Dm-Ftz között.
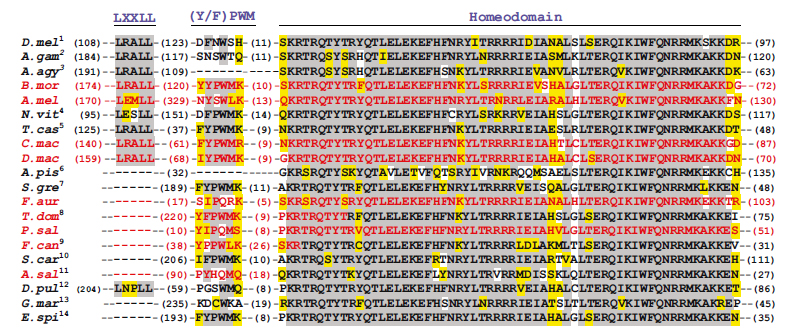
A mi szempontunkból a Dm-Ftz fehérje (a Drosophila meanogaster Fushi tarazu fehérjéje) két szakasza bír különös fontossággal: Az LXXLL motívum, ami lehetővé teszi, hogy komplexet képezzen a az Ftz-F1 fehérjével, ez szükséges a pair-rule működéshez. Az YPWM motívum pedig a homeotikus fehérjék működéséhez szükséges, ezzel képes komplexet képezni az Extradenticle nevű fehérjével, ez a Dm-Ftz fehérjében működésképtelen, vélhetőleg ezért nem működik HOX fehérjeként, bár a homeodoménje megvan és ugyanúgy képes kötni a DNS –t, mint a HOX fehérjék. A T. castaneum Ftz ortológja ép YPWM motívumot hordoz, nem meglepő módon homeotikus fehérjeként tud is működni. Ha a Dm-Ftz fehérjében visszaállították az YPWM motívumot, az is homeotikus fehérjeként működött. Ez a két rövid aminosavszekvencia változása felelős az Ftz fehérje működésének megváltozásáért.
Az ősi YPWM motívum már egészen más képet mutat. Mivel megtalálható a vizsgált homeotikus fehérjékben is (Antp, Scr), vélhetőleg az ősi Ftz fehérjében ez megvolt és később az evolúció során változott meg többször is, egymástól függetlenül. Azért nevezik őket degeneráltnak az ábrán, mert hasonlítanak ugyan a konszenzusszekvenciára, ám nem tűntek el teljesen, csak csökkent az Extredenticle kötő képességük. Azért gondolják, hogy minden ilyen esemény egyedi mutáció eredménye, mert minden egyes degenerált YPWM szekvencia másban tér el a konszenzusszekvenciától, ráadásul közeli rokon fajokban eltér a fehérje ezen szakasza.
E mellett megváltozott a Dm-Ftz kifejeződési mintázata is, egyetlen szelvényről hét pair-rule sávra. Ugyanezt a sávozott kifejeződési mintázatot mutatja a az előbb említett T. castaneum bogár Tc-Ftz fehérjéje és a mézelő méh Am-Ftz fehérjéje, ám például a Schistocerca gregaria nevű sáskában hiányzott ez a sávos kifejeződés. A gén kifejeződésében bekövetkezett változásokért nyilvánvalóan a szabályozóelemeinek a változásai felelősek. Hogy felrajzolhassák a Fushi tarazu fehérje evolúciós történetét, számos ma élő ízeltlábúban vizsgálták meg az Ftz fehérje működését, így húsz aminosavszekvenciából indult ki a vizsgálat. Mint az ábráról látszik az LXXLL motívum valahol a Holometabola csoport ősében alakulhatott ki, a vizsgált bogarakban, hártyásszárnyúakban és kétszárnyúakban is megtalálható, a többi ízeltlábúban viszont nem, a vízibolha nevezetes kivételével. Itt azonban sajnos nem állnak rendelkezésünkre funkcionális vizsgálatok, úgyhogy legföljebb találgatni lehet a szerepét illetően.
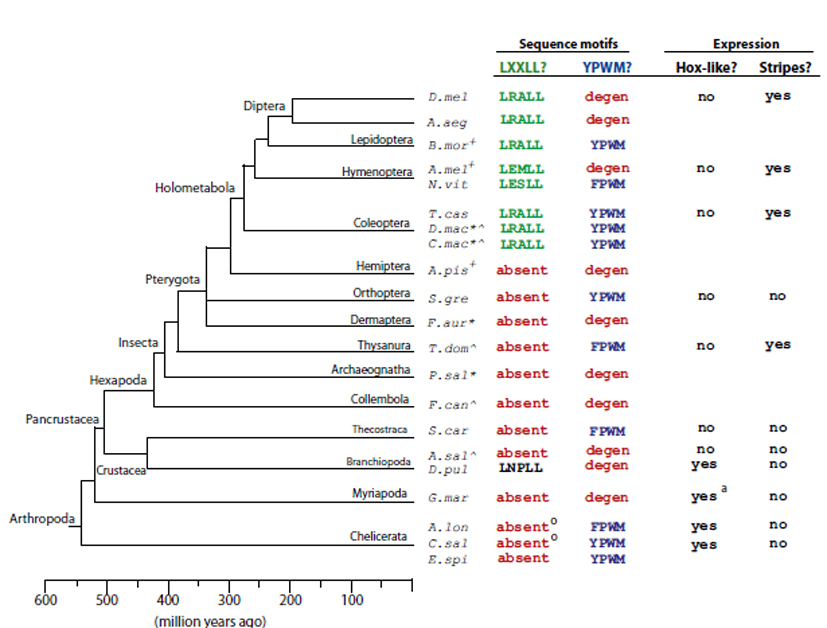
Ezek alapján fölrajzolható a Fushi tarazu fehérje evolúciós története: Egy ősi HOX gén duplikációjával létrejött a ma ismert Antp és Ftz fehérjék őse. Az ftz gén szabályozóelemei megváltoztak, lecsökkent az expressziója, így életképesek maradtak a különböző Ftz allélek, nem nehezedett rájuk olyan erős szelekciós nyomás. Ezek után kialakult benne az Ftz-F1 kötő motívum, így lehetővé vált, hogy szegmentációs fehérjeként működjön, majd beszerezte a szelvényezett kifejeződéshez szükséges szabályozóelemeket is, innentől már pair-rule génként működött. Ennek a folyamatnak az átmeneti állomásai ma is megtalálhatóak minden elképzelhető változatban, láthatóan életképes jószágokban.
Heffera A., Shultza J. W., Pick L. (2010): Surprising flexibility in a conserved Hox transcription factor over 550 million years of evolution; PNAS 2010 okt 4. (Epub ahead of print)
Löhrhr U.,Yussa M., Pick L. (2001): Drosophila fushi tarazu: a gene on the border of homeotic function; Current Biology 11:1403–1412
Sexcomb
Röviden azért már volt szó a ftz-ről, réges-régen ;-)):
http://criticalbiomass.freeblog.hu/archives/2006/01/11/Test_alkat_-_2/