
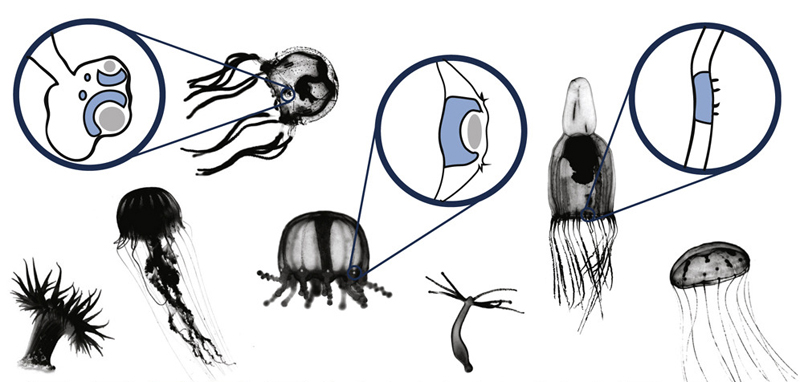
A szemevolúció mindig is hálás téma és egyike azon eseteknek, amikor a kreacionisták ignoranciából származó érvét (miszerint a komplex szem nem alakulhatott ki lépésenként) remekül lehet felhasználni pont annak a szemléltetésére, amit cáfolni szerettek volna.
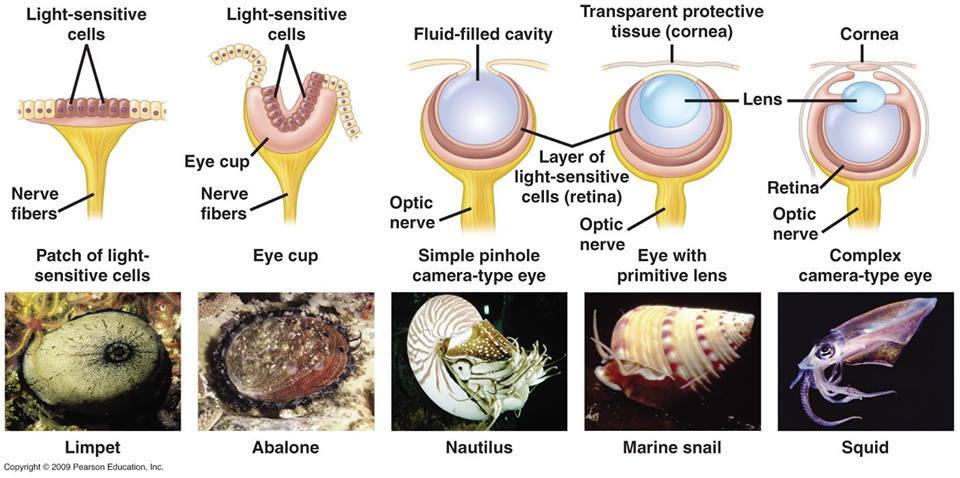
Szem, pontosabban fényérzékelő szerv sokszor kialakult az evolúció során. Elég csak a klasszikus, gerinces szem mellett a rovarok összetett szemére és a lábasfejűek más logikán működő kameraszemére gondolni, hogy belássuk ezek bizonyosan egymástól függetlenül jöttek lére – és ha egy csoporton belül jobban szétnézünk, akkor gyakran az is látható, hogy több, különböző logikájú és fejlettségű szem is megjelenhet, ami szintén azt sugallja, hogy a szem-szerű szervek evolúciója, ha már vannak fényérzékeny sejtek, viszonylag logikus kimenet.
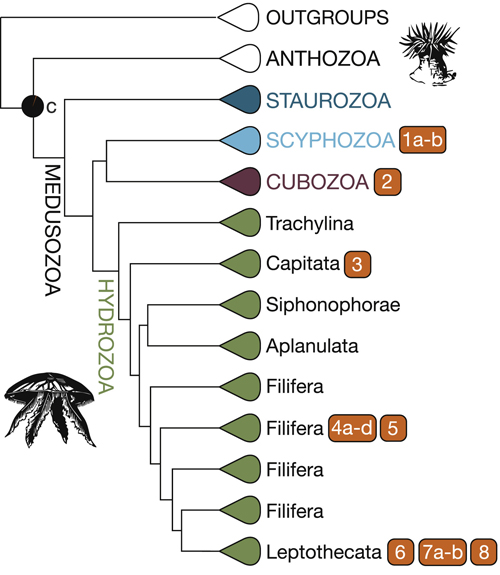
(Az alábbi összefoglaló ábrán az összetett szemeket oválisok, az egyetlen kamrával rendelkezőket pedig négyzetek jelölik. A színkódok elkülönítik az egyszerű, árnyékok érzékelésére alkalmas látószerveket (piros), a lencséket használóktól (kék) és a tükröződő felületekkel operálóktól (zöld).)
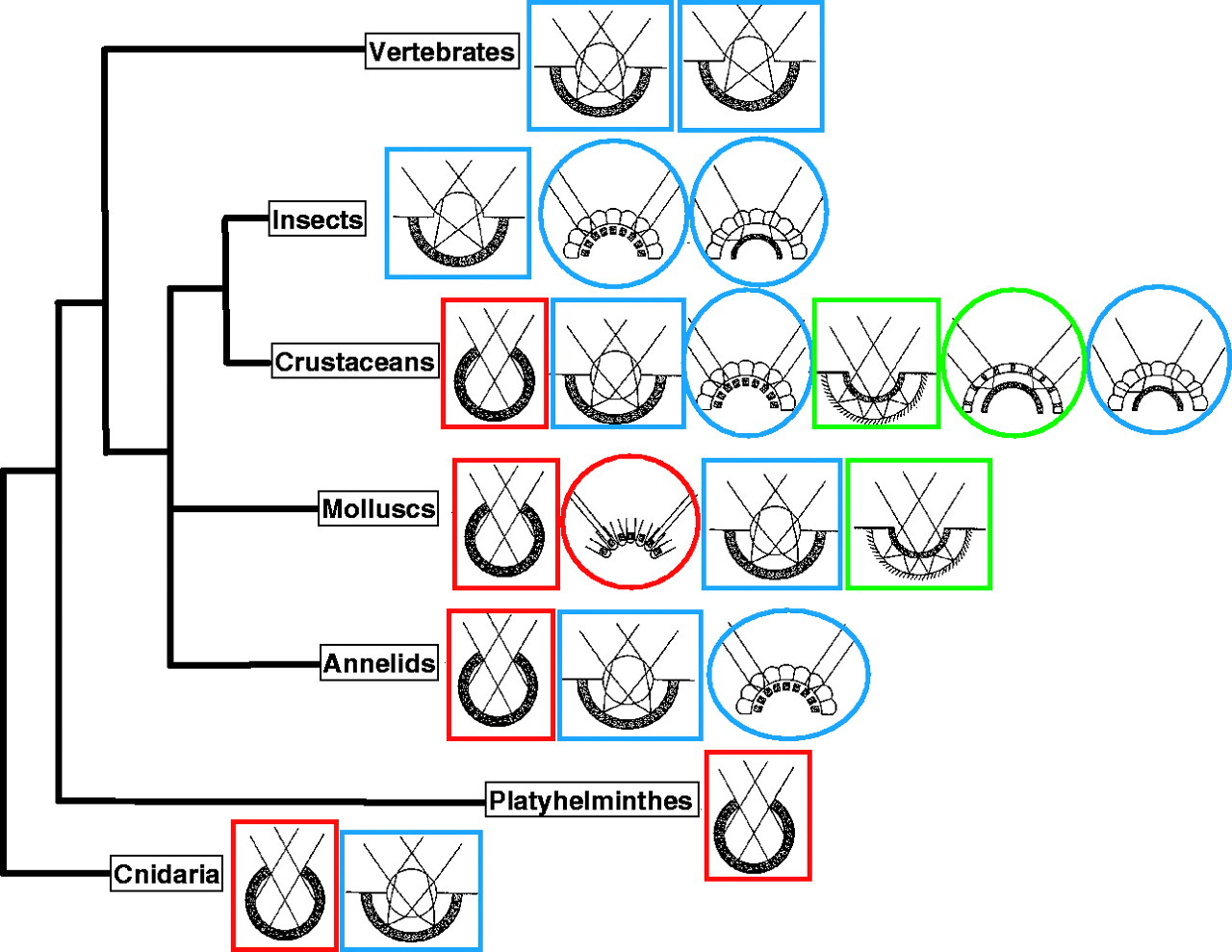
Akadékoskodók persze felvethetik, hogy ezek a szemek szintén nem jöhetnek egy lépésben létre – de természetesen ilyesmit nem is állít és például a puhatestűek esetében jól láthatók egyes fajokban azok az „átmeneti” struktúrák, amelyeket létezését már a korai, egyszerű számítógépes szimulációk is kiadták.
Pár hete a Current Biology-ban jelent meg egy tanulmány, ami az egyik legegyszerűbb felépítésű állatcsoport, a csalánzók esetében nézi meg molekuláris kladisztikát használva, hogy hányszor is alakulhattak ki „szemek”.
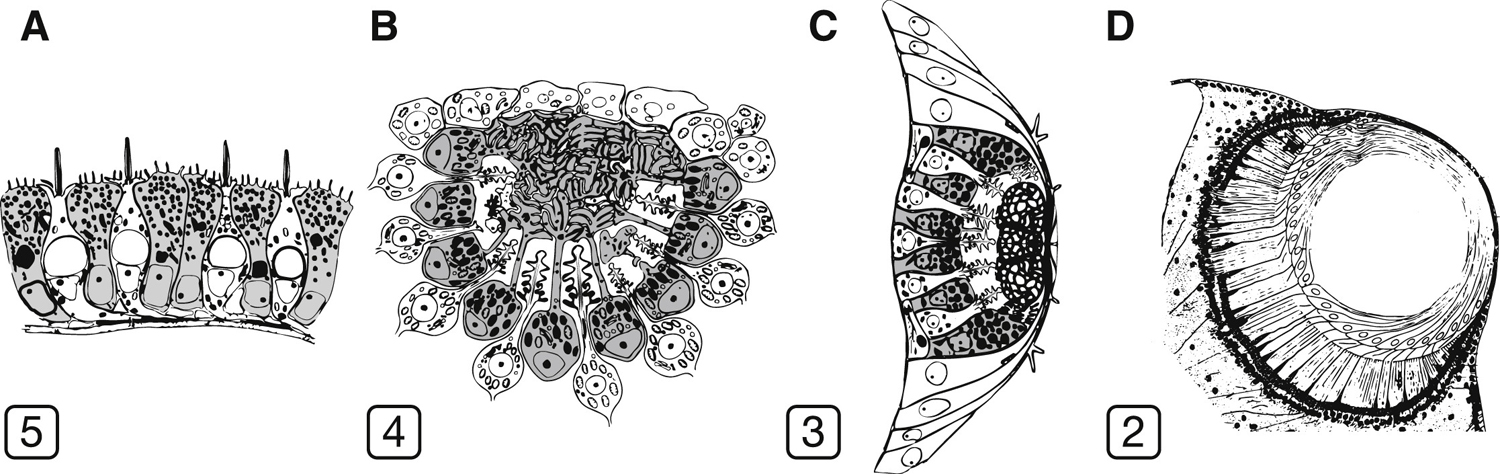
Az már az ultrastruktúrájuk alapján is valószínű volt, hogy ennyire különböző látószervek, mint az alábbi ábrán is láthatók (illetve röpke évtizede már írtam a kockamedúza remek szeméről is), egymásból nehezebben származtathatók, mint egy szem-mentes állapotból, de hogy ebben biztosak lehessünk, ahhoz kellett egy nagy pontosságú filogenetikai fa és azon rögtön jobban látszik, hogy sok „szemes” csoport közvetlen rokonságában szinte kizárólag szem nélküliek vannak.
Ez pedig már az egyszerű kladisztika szintjén is azt sugallná, hogy vagy a közös ősnek is volt valami szemre emlékeztető szerve, ami nagyon sokszor elvesződött az evolúció során, vagy többször – konkrétan nyolcszor – kialakult ebben az állatcsoportban is. A cikk szerzői ez utóbbi álláspont mellett teszik le voksukat és ennek alátámasztására megvizsgálják a különböző szemekben fellelhető opszinokat is – ti., ha a szemek újból és újból eltűntek volna, akkor a látószervvel rendelkezől szemében levő opszinok szorosabb rokonságot kellene mutassanak (monofiletikus csoportot alkotnának). De nem ezt látjuk, ami megintcsak azt támasztja alá, hogy a csalánozó szemek sokszor, egymástól függetlenül jöttek létre.
Ami persze nem teljesen véletlen, hiszen még a szemmel nem rendelkező fajok esetében is a bőrben számos fotoreceptív sejt van. Ezeket használta fel aztán az evolúció, hogy a korábban emlegetett szimulációknak megfelelően, újból és újból szemfoltokat, vagy annál is komplexebb struktúrákat hozzon létre.
Picciani N, Kerlin JR, Sierra N, Swafford AJM, Ramirez MD, et al. (2018) Prolific Origination of Eyes in Cnidaria with Co-option of Non-visual Opsins. Curr Biol 28(15): 2413-2419.e4
Treisman JE (2004) How to make an eye. Development 131(16): 3823-7
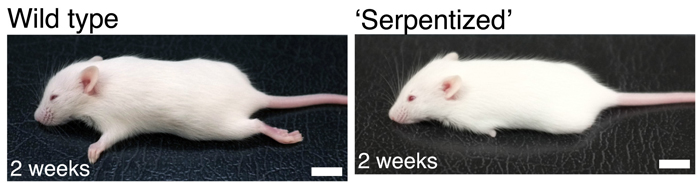


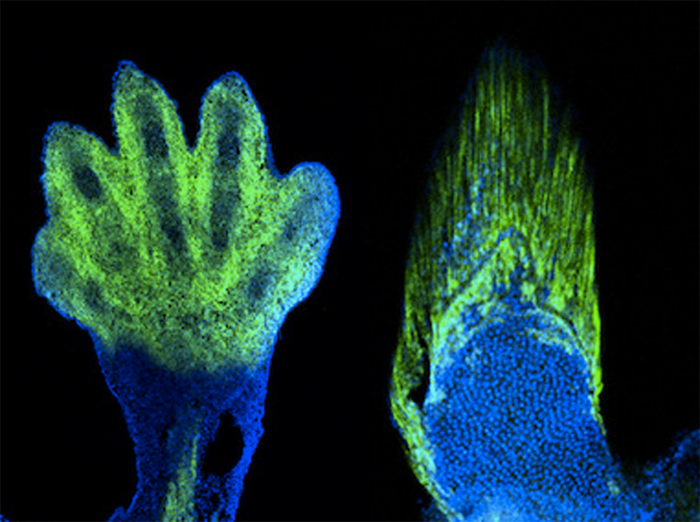

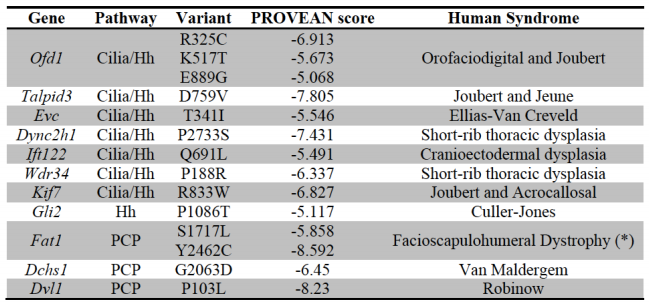
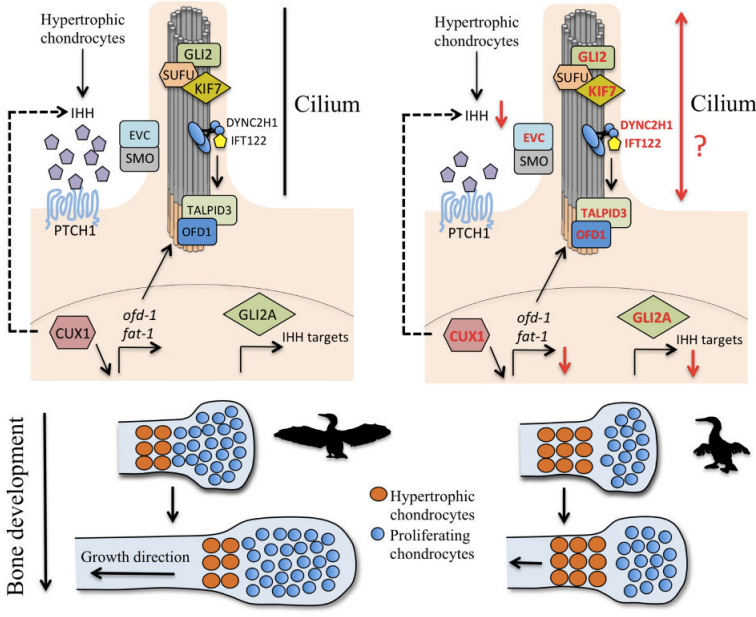
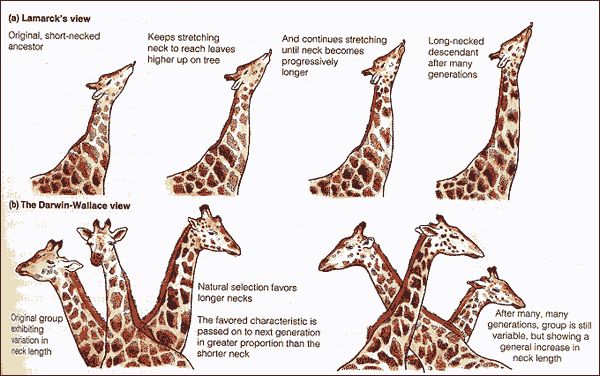 Ha van szó szerint tankönyvi példája a darwini evolúciónak, akkor az természetesen a zsiráfok kialakulása lenne. Hiszen ez az a folyamat, amivel leggyakrabban szemlélteni szokták a lamarcki és darwini evolúciós szemlélet közti különbségeket: míg a lamarcki nézet szerint a nyakát nyújtogató, okapi-szerű ős szorgalmas edzése vezetett a meghosszabbodott nyakhoz, addig a darwini nézet szerint a fajon belüli variancia azoknak az egyedeknek nyújtott szelekciós előnyt, akik hosszabb nyakkal rendelkeztek, ami hosszú generációk alatt az ő hosszú-nyakat biztosító génjeik elterjedését és így a faj/populáció általános nyakhosszabbodását okozta.
Ha van szó szerint tankönyvi példája a darwini evolúciónak, akkor az természetesen a zsiráfok kialakulása lenne. Hiszen ez az a folyamat, amivel leggyakrabban szemlélteni szokták a lamarcki és darwini evolúciós szemlélet közti különbségeket: míg a lamarcki nézet szerint a nyakát nyújtogató, okapi-szerű ős szorgalmas edzése vezetett a meghosszabbodott nyakhoz, addig a darwini nézet szerint a fajon belüli variancia azoknak az egyedeknek nyújtott szelekciós előnyt, akik hosszabb nyakkal rendelkeztek, ami hosszú generációk alatt az ő hosszú-nyakat biztosító génjeik elterjedését és így a faj/populáció általános nyakhosszabbodását okozta.
 Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.
Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.