A tavalyi hiátus után az idén valóban megrendezésre kerül az Ünnepi Könyvhét (még ha kis késéssel is). Mi pedig, szokásunkhoz híven erre az alkalommal egy külön könyvajánló poszttal készültünk, ami annyiban különleges, hogy először ajánlunk olyan könyvet, amelynek a szerzője egyben a blog (egykori és esetei) szerzője is. Lássuk hát, mit olvastak régi és új szerzőink az elmúlt évben, amit ajánlanának is a blog látogatóinak.
A homoszexualitás napjainkban egy megosztó téma. Országunkban jelenleg különösen aktuális kérdés: Elfogadható-e az azonos neműekkel folytatott intim kapcsolat, illetve szabad-e ennek létezéséről tudniuk a kiskorúaknak? Úgy tűnik, mostanság egyik sem kívánatos, köszönhetően egy új törvénymódosításnak. Noha ezt talán senkinek nem is kell részleteznem, a lényeget kiemelném.
…tilos tizennyolc éven aluliak számára pornográf, valamint olyan tartalmat elérhetővé tenni, amely a szexualitást öncélúan ábrázolja, illetve a születési nemnek megfelelő önazonosságtól való eltérést, a nem megváltoztatását, valamint a homoszexualitást népszerűsíti, jeleníti meg…
Javaslat módosítási szándék megfogalmazásához a T/16365. számú törvényjavaslathoz
Homoszexualitás és az emberi természet
A módosító hátterében minden bizonnyal az régóta hangoztatott érv áll, miszerint egy homoszexuális kapcsolat természetellenes (peccatum contra naturam, latin: természet ellen elkövetett bün), s ilyenektől védeni kell a fiatal generáció elméjét. De valóban természetellenes-e az azonos neműekkel folytatott viszony? Biológusként kicsit kötelességemnek érzem, hogy e témát pár szóban és pár példával körüljárjam.
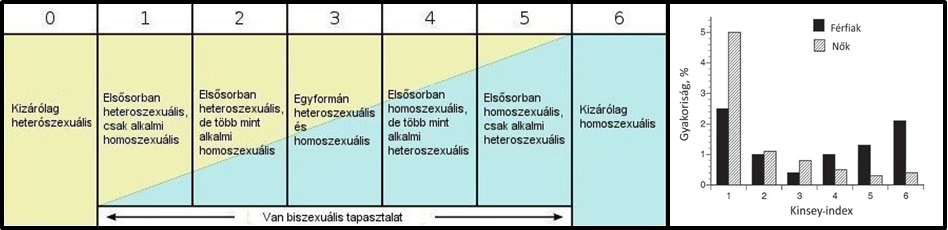
Nem fogok pontos definíciókkal élni. A következőkben a „homoszexualitás” alatt egyfajta „nem heteroszexuális” viselkedést fogok érteni. Kezdjük ott, hogy magát a homoszexualitást is nehéz kategorizálni. Manapság ez egyfajta kizárólagosságot hordoz magában. A témával való kutatások többsége azonban inkább egy skála (pl. Kinsey-skála) mentén szokta osztályozni a szexualitást attól függően, hogy valaki kizálólagosan preferál egy nemet, netán csak fantázia szintjén, esetleg alkalmilag érdeklődik az egyik vagy a másik iránt (1. ábra).
1. ábra: A Kinsey-skála, és a nem kizárólagosan heteroszexuális kategóriák eloszlása a nemek között. Megjegyzés:A kizárólagosan heteroszexuális csoportja (Kinsey 0) a népesség kb. 90%-át teszi ki, ez az ábrán nincs feltüntetve (Baley és mtsai. nyomán, forrás: Scheuring, 2014)
Például az ókori Görögországban és Rómában teljesen elfogadott volt házas férfiként, családapaként fiatal férfiú szeretőket tartani (bár Platón idejében indultak viták ennek „rendbontó” hatásairól). Más kultúrákban is komoly történelme van a homoszexualitásnak. Kínában már i.e. 600 körülről vannak feljegyzések homoszexuális viszonyokról, és ismeretesek egyéb, Közel- és Távol-Keleti példák is, mely nagyon fluid szexuális preferenciákról (vagy épp nemi szerepekről) számolnak be. E viszonyok morális megkérdőjelezése csak 1000-1200 évvel később, a keresztény és iszlám betörésével indul meg.
Szintén érdekes óceániai kultúrák koncepciója a nemi identitásról és szexuális preferenciáról. A polinéz nyelvekben nagyon régóta létezik szó „azonos neműek közötti intim kapcsolat”-ra (maori: takatāpui; hawaii: moe aikāne). James Cook legénységének beszámolói alapján ez teljesen elfogadott és stigma nélküli együttlét volt. Szintén érdekesség, hogy e népcsoportokban gyakran létezik a társadalmilag elfogadott, néha fontos kulturális szerepet betöltő „harmadik nem” megkülönböztetése (Hawaii: māhū; Cook szk.: ‘akava’ine; Tonga: fakaleitī; Tuvalu: pinapinaaine; Niue: fakafifine; Szamoa: faʻafafine). Ők férfiként saját nemükhöz vonzódnak, és számos tekintetben nőként viselkednek (egy széles spektrumban). Talán a transzneműség a legkifejezőbb szó, de nehéz lenne ezt teljes mértékben lefordítani a 21. századi LGBT paradigmára, és teljesen más kulturális identitás jellemzi például Szamoán a fa’afafinéket, akikről később még említést teszünk.
Tehát régóta léteznek kultúrák, ahol egyáltalán nem tabu az azonos neműek közötti kapcsolat. Vajon e népek elhajlása csak egy téves berögződés, netán valami sokkal általánosabb trend, mely túlmutat az emberi viselkedésen?
Homoszexualitás az állatvilágban?
Az utóbbi 20 év megfigyelései alapján ma már több mint 1500 faj esetében írtak már le homoszexuális viselkedést az állatvilágban (same-sex sexual activity: az egyed egy másik azonos nemű egyeddel létesít szexuális jellegű kapcsolatot). Akadnak köztük szép számmal rovarok, halak, kétéltűek, hüllők, madarak és emlősök is.
A házi juh (Ovis aries) esetében például a kosok egytizede elutasító nőstényekkel szemben, míg más kosokkal előszeretettel kezdenek ki. A Laysan albatrosz (Phoebastria immutabilis) Oahu szigetén fészkelő populációjánál a fészkelő párok kb. egyharmada nőstény-nőstény pár. Egyes megfigyelések alapján a zsiráfbikák is gyakrabban udvarolnak egymásnak mint nőstényeknek. És a főemlősök sem képeznek kivételt: japán makákók, gorillák és orángutánok esetében is leírtak már azonos nemű közösüléseket.
Az ember egyik legközelebbi rokona, a törpecsimpánzok (Pan paniscus) példája maga a szodomizmus netovábbja. Nemcsak, hogy nagyon is „öncélúan” élnek a szexuális aktivitással, hanem itt a biszexualitás a norma, az aktusok több mint fele kettő vagy több nőstény között történik. De a hímek közötti szex is bevett szokás! Társadalmukban a szex sokkalta több szereppel bír mint az utódnemzés funkciója. De erre majd később visszatérünk.
2. ábra: Néhány állatfaj, jól dokumentált homoszexuális viselkedéssel: közönséges házi juh, zsiráf hím-hím pár, Laysan albatrosz nősténypár, és a bizonyítottan biszexuális bonobók bájosan bámészkodó bandája. (forrás: wikipedia.com)
És ez csak pár kiragadott példa volt. Tehát közel sem állítható az, hogy a homoszexualitás pusztán valami „mentális perverzió” lenne. Ez egy viszonylag gyakori, sőt mondjuk ki, ez igenis természetes és gyakran előforduló viselkedési forma az állatvilágban is.
Isten és Darwin nevében
Akkor miért tarthatjuk természetellenesnek e viselkedést? Ha amúgy az állatvilágban gyakori stratégia, akkor lehet hogy pont a kulturális hátterünk miatt? Nem szeretnék valakinek a hitvilágába belegázolni s azt a végletekig boncolni, de azért spekulálhatunk (ilyenek ezek a spekulánsok!), mint egy evolúcióbiológus. Innen kapargatva a felszínt talán jobban érthető lesz, hogy miért tapasztalható meg egyfajta értetlenség és mélyen gyökerező ellenszenv a homoszexualitással szemben.
Valószínűsithető, hogy a homoszexualitás morális megkérdőjelezése főképp az ábrahámi vallásoktól (pl.: zsidó, keresztény, iszlám) és azok későbbi befolyásából (nyugatiosodásból, kolonizáció) ered. Ironikus módon ugyanis e világvallások értékrendje meglepően „darwinista”, legalábbis tartogat olyan alapvetéseket mely egy evolúcióbiológus fantáziáját is megmozgatja.
E vallások egyfajta adaptív funkciója, hogy egy összetartóbb és erősebb humán csoport megszervezésére törekszik egy „kulturális kódex” segítségével. Az egyik ilyen érdekes aspektus például a Nagy Testvér személye (nevezzük bármilyen néven is): az emberek többsége sokkalta jobban kooperál (vagy mellőzi a csalásokat bizonyos szituációkban) ha van egy külső megfigyelő, hisz az maga után vonhat egy későbbi megtorlást. Márpedig egy omnipotens entitás mindent lát és minden kihágást büntethet.
Egy másik ilyen, és mostani témánk szempontjából fontosabb, dogma a szex társadalmi funkciójának tisztázása a csoport érdekében. Ha különböző csoportok versengenek, akkor fontos, hogy melyik csoport szaporodik gyorsabban. Az lesz előnyben a versenytársakkal szemben. A feltételezés esetünkben pedig az, hogy nagyobb szaporodási rátát lehet elérni azzal, ha mindenféle „öncélú szexuális tevékenység” helyett a ráfordítható energia egy családba, illetve egy exkluzív partnerrel való utódgondozásba megy. Részben ezért is válik természetellenessé és elítéltté egy homoszexuális kapcsolat: nincs belőle utód, a csoport számára pedig ez nem feltétlen előnyös.
A darwini paradoxon és lehetséges megoldások
De nem csak társadalmi, hanem evolúciobiológiai szempontból is paradoxonnak tűnhet az azonos neműekkel folytatott viszony. Valószínűleg van a viselkedésnek egy öröklődő, genetikai háttere. Scheuring István előadása és korábbi Természet Világa cikke ezt sokkal élvezetesebben vesézi ki, csak ajánlani tudom (és nem tagadnám, hogy nagy hatással voltak e posztra is). Viszont ha ez örökletes, és homoszexuális embereknek kevesebb utódja születik (nem feltétlen nulla), akkor a „homoszexuális” génnek ki kellene szelektálódnia. De nem ezt látjuk. Ez pedig egy érdekes evolúciós kérdés: Miért nem tűnik el a homoszexuális viselkedés a populációkból?
Lehet, hogy a homoszexualitásnak van valamilyen közvetett előnye? A Laysan albatrosz esetében a kiváltó ok a hímek hiánya: a nőstény-nőstény párok költési sikere noha kicsi (az apa ilyenkor egy alkalmilag félrelépő hím), de a sikeres költés esetén több esélyük van a következő idényben hím párt fogni maguknak. Egyes, hímeket teljesen nélkülöző és önmaguk megtermékenyítésére képes (ún. partenogenetikus) gyíkfajoknál (pl. Aspidoscelis neomexicanus) a nőstények közötti szex valószínűleg elengedhetetlen az önmegtermékenyítés a beindításához.
Az ember esetében rokonszelekció gyanúja merül fel mint lehetséges mechanizmus. Ha például egy homoszexuális egyed utódnevelés helyett inkább a rokonait segíti, azzal továbbra is hozzájárulhat a hajlamosító gén terjedéséhez. Az ugyanis a rokonban is nagy eséllyel megtalálható. Pusztán arra van szükség, hogy a segítség mértéke és a rokonsági fok meghaladja a homoszexuális egyed költségét. Ez a közvetlen előny már fenntarthatja a homoszexualitást a „hordozók” segítésével.
Vannak eredmények mely ezt a rokonszelekciós narratívát erősíthetik, bár korántsem beszélhetünk átfogó bizonyítékokról. Több kutatás (például ez is) kimutatta, hogy heteroszexuálisokhoz képest a homoszexuálisok nőrokonainak magasabb az átlagos utódszáma; ez egy szaporodásbeli előny, és X-kromoszómához kötött öröklődésre is utal. Azt sajnos nem sikerült kideríteni, hogy ez a női rokonok felé nyújtott segítség pontosan miben jelenik meg.
Talán ebből a szempontból érdekesek a korábban már említett szamoai fa’afafine-k (ők a már említett harmadik nem tagjai, nőies férfiak akik férfiakhoz vonzódnak). Esetükben kimutathatő, hogy sokkal jobban aggódnak a testvéreikért mint a heteroszexuálisok. Döntési kényszerek esetén is jobban preferálják azon rokoni kategóriák támogatását, amely a genetikai rokonságot és a rokoni előnyt maximalizálja. Ezek a kategóriák éppenséggel a lánytestvérek, azok gyermekei, és ott is elsősorban a lányok. És hogy miért a nők? Az anya kiléte sosem kérdés, így rokonság mindig biztosabb lesz a női vonalon.
Ugyanakkor érdemes megemlíteni, hogy a fa’afafine-k esetében talált eredmények a rájuk jellemző nőies viselkedéssel és a társadalmi háttérrel is összefüggésbe hozhatók, nem feltétlen csak a homoszexualitással. Nyugati társadalmak és Japán homoszexuális férfiainál például nem figyelhetők meg ilyen döntési trendek. Tehát elképzelhetők más, genetikai és nem rokonszelekción alapuló mechanizmusok is amik fenntartják a homoszexualitást.
És ha már társadalom, szóba jönnek a törpecsimpánzok. Náluk a főképp alkalmi, homoszexuális kapcsolatoknak elsősorban feszültségcsökkentő és békítő szerepe van. Ez a társadalmi csillapító szerep hozzájárulhat a stabil közösségek megszervezéséhez. Tekintve, hogy az ember egyik legközelebbi rokonáról van szó, felmerül a kérdés: emberelődöknél, netán korábban emlegetett emberi kultúráknál ez mekkora szerepet játszhatott az azonos neműek közötti kapcsolat a nagyobb egyedcsoportok, társadalmak kialakításában?
Zárszó
A sokféleség Földünk és annak élővilágának velejárója. Minden fajon belül találunk kisebb és nagyobb egyedeket, színváltozatokat. A szexuális preferencia is csak egy ilyen dolog, nem egy természetellenes viselkedés. Érthető, hogy társadalmi közegünkben miért tűnik értelmetlennek, azonban ha jobban utánajárunk, rájöhetünk, hogy egyrészt a homoszexualitás nem ritka, másrészt akadnak logikus magyarázatok létezésére.
És itt szó sincs népszerűsítésről. Ez egy jelenség, a létezését tudomásul kell venni, pláne ha az továbbra is hátrányos megkülönböztetéshez vezet. Nem népszerűsíteni kell, nem is kell félni tőle, hanem megérteni. Ha valaki engem (mint evolúcióbiológust) kérdez, a fiatalabb generációt megakadályozni a világ és a sokféleség megértésében igenis hátrányos lehet. Pláne ha egy jelenségnek megvan a helye a nemcsak a természetben, de társadalmunkban is.
Scheuring, I. (2014) ‘A homoszexualitás evolúciógenetikai háttere’ Természet Világa, 145 (11). pp. 496-500.
Bailey, J. M., Dunne, M. P. and Martin, N. G. (2000) ‘Genetic and environmental influences on sexual orientation and its correlates in an Australian twin sample.’, Journal of Personality and Social Psychology, 78(3), pp. 524–536. doi: 10.1037/0022-3514.78.3.524.
Iemmola, F. and Camperio Ciani, A. (2009) ‘New Evidence of Genetic Factors Influencing Sexual Orientation in Men: Female Fecundity Increase in the Maternal Line’, Archives of Sexual Behavior, 38(3), pp. 393–399. doi: 10.1007/s10508-008-9381-6.
VanderLaan, D. P., Petterson, L. J. and Vasey, P. L. (2017) ‘Elevated Kin-Directed Altruism Emerges in Childhood and Is Linked to Feminine Gender Expression in Samoan Fa’afafine: A Retrospective Study’, Archives of Sexual Behavior, 46(1), pp. 95–108. doi: 10.1007/s10508-016-0884-2.
VanderLaan, D. P. and Vasey, P. L. (2014) ‘Evidence of Cognitive Biases for Maximizing Indirect Fitness in Samoan Fa’afafine’, Archives of Sexual Behavior, 43(5), pp. 1009–1022. doi: 10.1007/s10508-014-0288-0.
De Waal, F. B. M. (2006) ‘Bonobo Sex and Society’, Scientific American, 16(2), pp. 14–21. doi: 10.1038/scientificamerican0606-14sp.
Wilson, D. S. (2002) Darwin’s Cathedral. University of Chicago Press. doi: 10.7208/chicago/9780226901374.001.0001.
Young, L. C. and VanderWerf, E. A. (2014) ‘Adaptive value of same-sex pairing in Laysan albatross’, Proceedings of the Royal Society B: Biological Sciences, 281(1775), p. 20132473. doi: 10.1098/rspb.2013.2473.
A napokban jelent meg egy régóta dédelgetett, Kun Ádám-mal közösen írt cikkünk a Scientific Reports-ban. Gondoltam írok róla egy „akadémiai hokedlifoglalót”, hátha elő tudom adni azt érdekesen. Még akár akkor is, ha az olvasó nem biológus. Szemtelen önmarketing következik…
A genetikai kód és a QWERTY billentyűzet talányai
Akad pár hasonlóság az élő szervezetek és civilizációnk vívmányai között. Én napjaimat azzal töltöm, hogy betűk formájában vetem papírra a fejemben keringő gondolatokat. Ehhez a billentyűzet megfelelő gombjait kell lenyomnom egymás után. Ezekből a betűkből végül szavak, és mondatok lesznek.
Ugyanilyen beleéléssel püföli saját kis genetikai billentyűzetét a Föld összes organizmusa, hogy a DNS-ben található, kódolt információt lefordítsa az életfolyamatokhoz nélkülözhetetlen fehérjékre. Ebben az esetben a DNS-en hordozott bázishármasok (pl. AGC, CAA) határoznak meg egy aminosav-sorrendet, mely a fehérjéket alkotja.
A hasonlóság itt nem ér véget. Az emberek túlnyomó többsége ugyanis majdnem ugyanazt a „QWERTY” billentyűkiosztást, vagy annak variánsát használja (1. ábra; a magyar QWERTZ billentyűzet ezzel szinte teljes mértékben megegyezik). Ennek egyik következménye, hogy ékezetektől eltekintve vakon képesek lennénk ugyanazt a szöveget bepötyögni bárhol. Az adott karakterek pozíciója nem változik a klaviatúrán, legyen szó számítógépről vagy telefononról, Svédországtól egészen Kínáig.
1. ábra: A világszerte használt QWERTY billentyűkiosztás (forrás: wikipedia.hu)
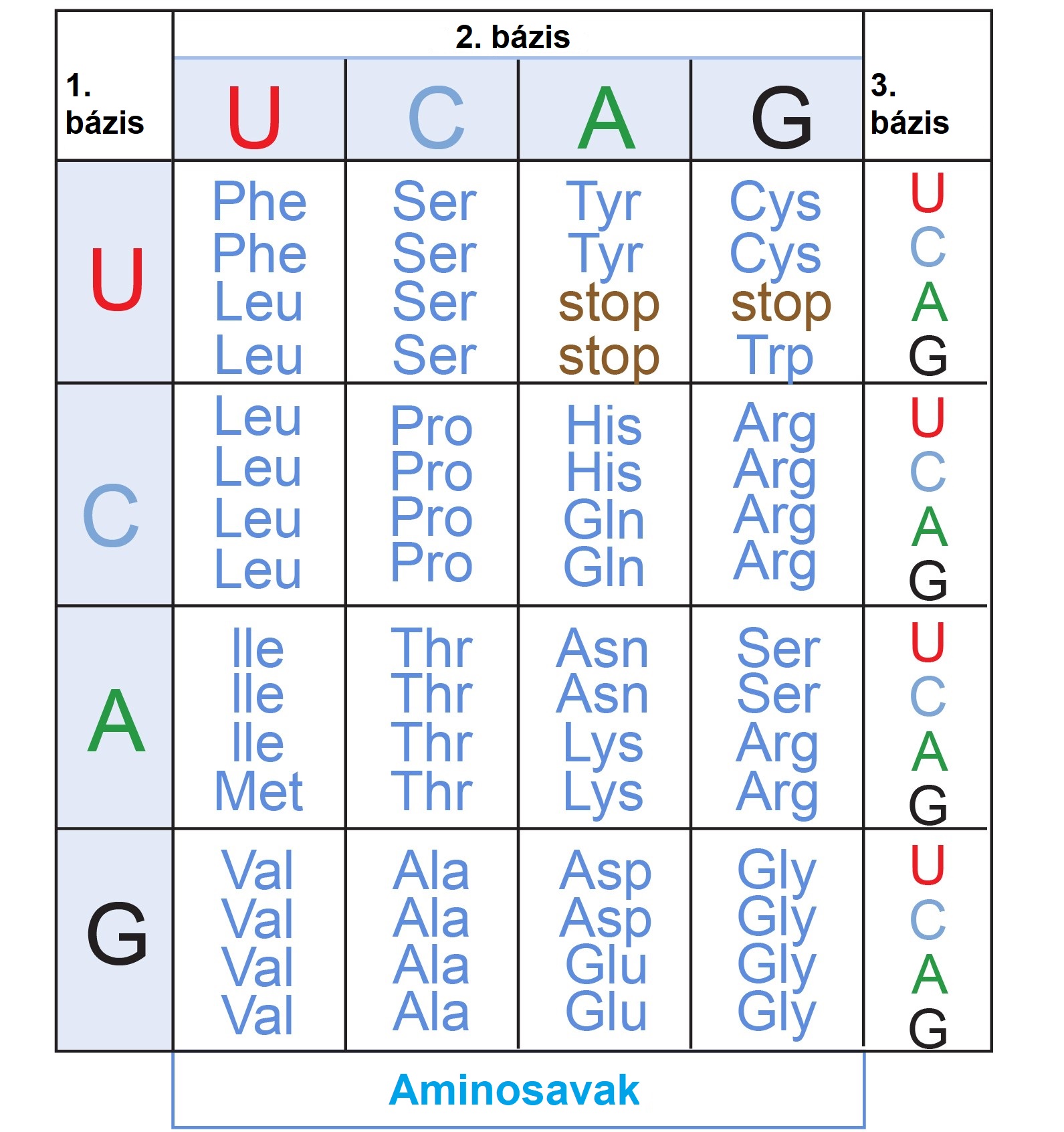
Ehhez hasonlóan minden organizmusban ugyanaz a genetikai ”billentyűkiosztás” található meg! Ha fogok egy emberi DNS-darabot, és azt egy másik élőlénybe, például egy baktériumba ültetem át, megkaphatjuk ugyanazt az fehérjét, ami bennem is termelődik. Egy adott bázishármas, csakúgy mint egy leütés helye a klaviatúrán, ugyanarra a karakterre (aminosavra), fog dekódolódni, pl. CCA → prolin, AGG → arginin (2. ábra). Mindenkiben ugyanaz a fordítócsomag, ugyanaz a genetikai kód fut.
2. ábra: A standard genetikai kód, az élő szervezetek billentyűkiosztása
Hiába hasonlít a két példa csak felületesen egymásra, mindkettő felvet egy közös kérdést:
Miért pont ezek a kiosztások terjedtek el?
Miért használ mindenki QWERTY billentyűzetet és miért pont ez standard genetikai kód található meg a Föld valamennyi élőlényében? Milliárdnyi más felosztást el tudunk képzelni. Például a billentyűzeten valaki igazán sorba is rendezhette volna a betűket, mégse azt használjuk. Hasonlóan máig nem tudjuk, hogy miért pont ez egy genetikai billentyűzet terjedt el a Földön és ez miként alakult ki több milliárd évvel ezelőtt. Ez a Földi élet keletkezésének egy kulcskérdése. A kirakós egyik darabkájának feltárásában lesz segítségünkre ha megfejtjük az emberi billentyűzetet.
Miért QWERTY?
Magát a QWERTY-kiosztást még az 1870-es években fejlesztette ki Christopher Latham Sholes. Tervezése a kényelmes, lehető leghatékonyabb gépeléshez lett igazítva. Ehhez a legyakoribb karaktereknek és karakterkombinációknak könnyen kézre kell esniük, s ez az ist jelenti hogy gépelés során az egymást követő betűk leütéséhez a lehető legkisebb távolságot kell megtennie az ujjaknak, mindezt a lehető leggyorsabban.
Ugyanakkor a karakterek gyakorisága függ a beszélt nyelvtől is! Jelentős eltéréseket találhatunk az egyes nyelvek között (3. ábra). Például a kínai pinjin (a mandarin latin betűs átirata) nagyon elüt az európai nyelvektől ebben a tekintetben. Nem mindegy tehát hogy milyen nyelven gépelünk a QWERTY billentyűzeten.
3. ábra: Betűgyakoriságok különböző nyelvek írott formájában (forrás: Xiaojun Bi és mtsai., 2012)
Ezek a különbségek kihathatnak a gépelés hatékonyságára is. Ezt akár mi is könnyűszerrel ellenőrizhetjük (gép előtt ülők előnyben). Az angolban a leggyakoribb egymást követő karakterpárok: t-h, h-g, i-o, míg a pinjinben: z-h, c-h, s-h. Csak nézzük meg mekkorák a távolságok az egyes billentyűk között!
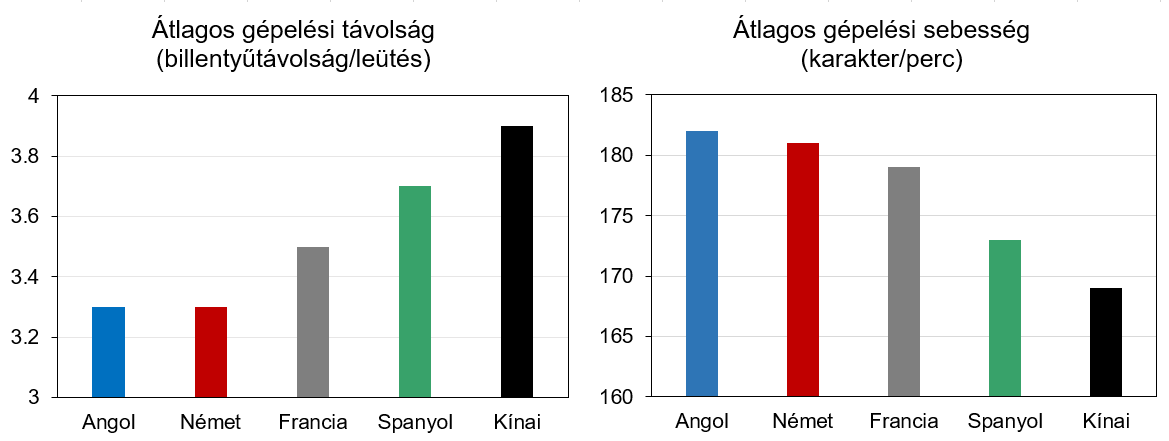
De ha ez nem lenne elég, akár tovább is mehetünk. Kiszámolható, és kísérletesen ellenőrizhető ugyanis, hogy milyen hatékony a QWERTY billentyűzet az egyes nyelvek esetén (4. ábra). Jól látható, hogy az billentyűk közti távolság az angolban és a hozzá valamelyest hasonlító németben a legkisebb. Őket követi a francia és a spanyol. A kínai pinjin-ben sokkal nagyobb távolságokat kell megtenni leütések közben, ez pedig azt is eredményezi hogy ezen a nyelven sokkal lassabban lehet csak gépelni.
4. ábra: Gépelési hatékonyságok különböző nyelveken, a QWERTY billentyűzetet használva (adatok: Xiaojun Bi és mtsai., 2012)
Az okot valószínűleg már az olvasó is gyanítja: e különbségek elsődleges oka az, hogy a QWERTY-kiosztást amerikaiak tervezték, az angol nyelv igényeire szabva. Minél kevésbé hasonlít egy nyelv az angolra, az annál inkább küszködni fog ezzel az univerzális billentyűzettel. A mi kutatásunk egy hasonló gondolatmenetet követett a genetikai kód eredetének feltárásához.
Mit tudtunk meg a genetikai kódról?
Cikkünk egy hasonló jelenség megfigyelése volt a genetikai kóddal kapcsolatban. Azt már sejtjük egy ideje, hogy a genetikai kód valamelyest optimalizálódott a fehérjék gyártásához. Ennek vannak egyértelmű jelei. Például régóta ismert, hogy a genetikai kód próbálja csökkenteni a „félreütés” kockázatát: ha történik egy mutáció a DNS-ben, az továbbra is ugyanazt, vagy hasonló tulajdonságú aminosavat fog eredményezni a fehérjében. Ezzel pedig lesz rá esély, hogy a „félregépelt” fehérje továbbra is működőképes marad.
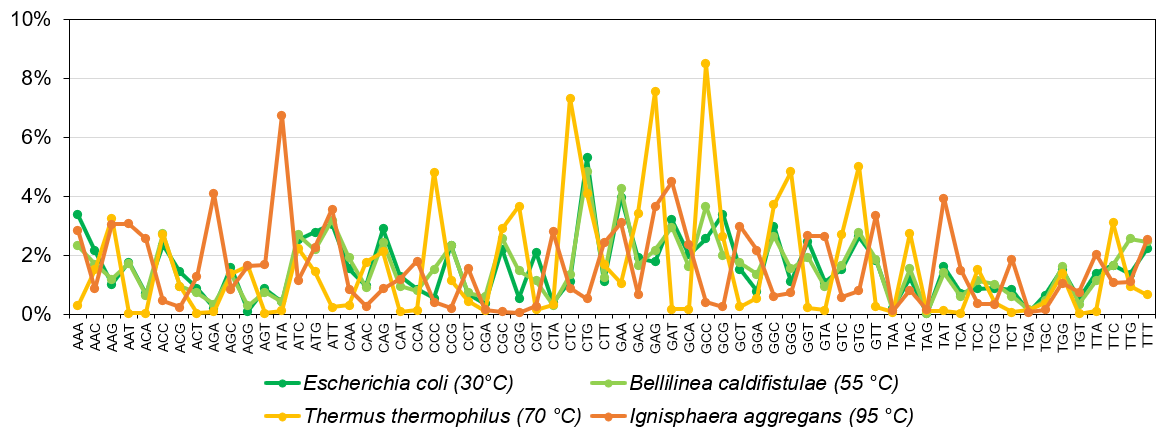
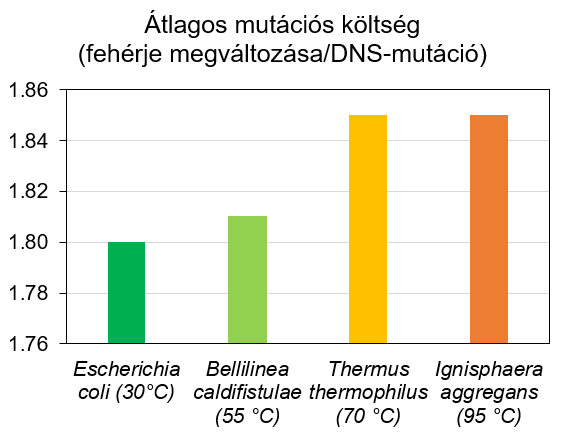
Az egységes kód ellenére azonban az élőlények is különböző „nyelvjárásokat” használhatnak, más gyakorisággal püfölik e genetikai billentyűzet gombjait (5. ábra). Az eltérés részben a környezet függvénye. Egy bélbaktérium (Escherichia coli) más összetételű fehérjéket igényel, mint pokoli körülményeket is tűrő mikrobák, akik akár még forráspont körüli termálvizekben is vidáman lubickolnak (pl.: Thermus thermophilus és Ignisphaera aggregans).
5. ábra: Bázishármas-gyakoriságok különböző mikrobák fehérjekódoló DNS-ében
Noha a különbségek nem nagyobbak mint irodalmi magyar és a szögedi ömbörökböszéde közti eltérés, az mégis kihathat genetikai kód hatékonyságára. Mi többek között azt mutattuk ki, hogy a forróbb élőhelyeket meghódító organizmusok e téren igenis alulmaradnak a hűvösebb környezetekben éldegélő „szobabacikkal” szemben (6. ábra). A standard genetikai kódot használva nagyobb a mutációk költsége az ún. extremofil organizmusokban, s így ők nem tudnak olyan hatékonyan gépelni.
6. ábra: A genetikai kód hatékonysága különböző mikrobák és azok optimális környezeti hőmérséklete esetén
Miért meglepő ez?
Mindez akár azt is jelenheti, hogy a genetikai kód jelenlegi formája nem biztos hogy extrém forró körülményekre lett kifejlesztve. Sokkal inkább egy enyhe, tavaszias időjárás képe bontakozik ki. Ez részben meghökkentő. Előszeretettel szeretjük azt gondolni, hogy hajdanán az Élet valami Yellowstone Nemzeti Park-szerű világban látott napvilágot, netán mélyvízi vulkanikus füstölgőkből kúszott elő. Én személy szerint mindig úgy képzeltem el az Élet bölcsőjét mint Csodaturmix varázsfőzetét forrni az üstben.
Azonban korántsem lehetetlen az sem, hogy valamikor hűvösebb körülmények uralkodtak a Földön, és ez adott otthont az első mikroorganizmusoknak. Az viszont, hogy ennek forgatókönyvnek a jelentőségét tisztázzuk és komolyan alátámaszthassunk, már a jövő zenéje…
És természetesen ez nem jelenti azt, hogy sikerült minden kérdésre választ találni a genetikai kóddal kapcsolatban. De lett egy új puzzle darabkánk, aminek segítségével több esélyünk lesz kirakni egy teljesebb képet az Élet keletkezésének korai fázisairól.
Források:
Radványi Á. & Kun Á. (2021) Phylogenetic analysis of mutational robustness based on codon usage supports that the standard genetic code does not prefer extreme environments. Sci Rep11, 10963.
Bi X., Smith B. A. & Zhai S. (2012) Multilingual touchscreen keyboard design and optimization. Human–Computer Interact27, 352–382.
Egy kellemes tavaszi délutánon az erdőszélen sétálva jó eséllyel botlunk lomha, nagy potrohú, fémesen csillogó kis drágakövekbe, amint hosszú, vézna lábaikkal esetlenül vonszolják testüket a talajon. Ők a hólyaghúzófélék családjába tartozó, és sokak által jól ismert nünükék (Meloesp.). A nemzetségnek hazánkban hozzávetőleg egy tucatnyi faja él, és mindegyikükre jellemző, hogy lárvakorukban önjáró kakukkfiókaként anyjuknál sokkal fürgébb és gondoskodóbb nevelőszülőkre kényszerítik rá magukat. Egészen konkrétan méhekre.
A nőstény nünükék a peték lerakását követően röviddel elpusztulnak (ez plusz egy érv a nevelőszülők keresése mellett). A petékből tavasszal, vagy nyár elején kikelő háromkarmú (triungulinus) lárvák felkapaszkodnak a virágzó növényekre, és a porzószálak közt meglapulva várnak a gyanútlan beporzókra. Karmocskáikkal aztán az arra szálló méhek kitinszőreibe kapaszkodnak, és a fészekbe vitetik magukat. A célpontok jellemzően magányosan élő (szoliter) méhfajok, mint az Andrena, Anthophora és a Chalicodoma nemzetségek tagjai. A triungulinus lárva a méhsejtbe kerülve először az ott lévő tojást fogyasztja el, majd egy másik lárvastádiumot öltve a felhalmozott mézkészleteket kezdi felfalni. Mindeközben egy relatíve biztonságos helyen fejlődhet. Lenyűgöző és mindenekelőtt kényelmes életstratégia ez, a kleptoparazitizmus (más fajok zsákmányának, illetve önfenntartás vagy utódgondozás céljából felhalmozott készleteinek megdézsmálása) egy érdekes alesete.
Európai rokonainál egy fokkal erőszakosabb módon érnek célba egy amerikai nünükefaj, a Meloe franciscanus lárvái. Ez a faj az Egyesült Államok nyugati területeinek félsivatagos, sivatagos élőhelyein érzi magát igazán otthon. Egy-egy magasabb kórón a triungulinus lárvák egész kis tömege szokott összeverődni, egy magányos méhfaj nőstény egyedének sziluettjét és illatát utánozva. Ilyen magasszintű kooperációra euszociális rovarok körében bőséggel akad példa, de bogaraknál még csak hasonlót sem figyeltek meg eddig. Főleg egzotikus ez a viselkedési forma, mivel egy speciális mimikrivel is társul.
Egy szerelemtől vak, és párzásra kész hím méhet, ha sikerrel megtéveszt a Meloe franciscanus lárvák nyüzsgő kupacának látványa, és a lárvák által termelt feromonfelhő, amaz a párzás reményében túl közel merészkedik a lárvakupachoz, és ezzel kész is a baj. A lárvák megrohamozzák a méhet, és amennyien csak tudnak, belekapaszkodnak a bundájába. A testsúlyeloszlás hirtelen megváltozása miatt a lépre csalt méh gyakran drámai módon a földre zuhan, majd miután a lárvák kényelmesen elhelyezkednek rajta, és ő visszanyeri egyensúlyát tovább repül, hogy ezúttal egy valódi nőstényre akadjon. Miután méhünk sikeresen rátalált szíve választottjára, és beteljesült szerelmük, a nünükelárvák, mint valami nemi betegség, átragadnak a nőstényre. Innentől kezdve az ő bundáján laknak, és türelmesen várják, hogy a nőstény megépítse a fészket, lerakja petéit és megkezdje a méz felhalmozását. Ekkor aztán eljön a triungulinusok ideje! Belevetik magukat a mézbe, s innentől kezdve európai unokatestvéreikhez hasonló módon tobzódnak a jólétben.
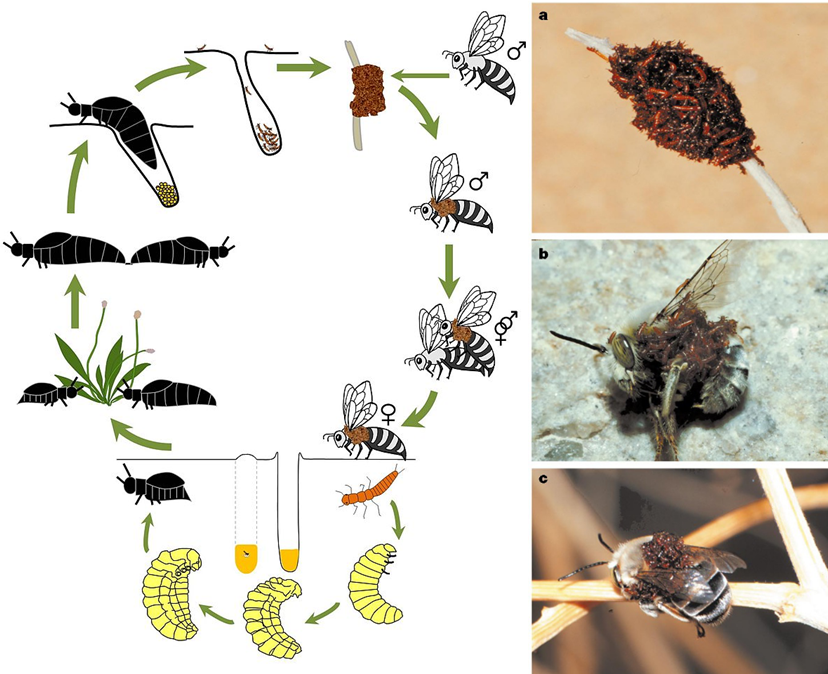
A Meloe franciscanus életciklusa (bal) (forrás: Wikipedia), triungulinus lárvák kupaca egy elszáradt növény szárán (a) és a megtévesztett méh testén (b; c) (forrás: Hafernik és Saul-Gershenz).
Belátható, hogy a M. franciscanus sikere azon áll, vagy bukik, hogy a lárvák magukhoz tudják-e vonzani a hím méhet. Minthogy ezt elsősorban az adott méhfaj nőstényeire jellemző feromonok termelésével érik el, a kleptoparazitizmus itt erősen gazdaspecifikus. A méhek ha szabadulni akarnak a nünükéktől, egyetlen evolúciós út járható számukra: változtatni kell egy kicsit a faj szexferomonjainak kémiai sajátosságain. Ez azonban könnyen reproduktív izolációhoz vezet az eredeti méhfajon belül, mivel az egyik populáció hímjei előbb utóbb nem lesznek képesek érzékelni egy másik populációba tartozó nőstény illatát. A populációk tehát előbb-utóbb faji szinten is elválnak egymástól.
Persze a parazitának is lépést kell tartania a „feromonkódok” újraírásával, ha nem akar gazda nélkül maradni. Erre szépen rímel az a megfigyelés, hogy a M. franciscanus Oregon part menti dűnéin élő populációja a Habropodamiserabilis nevű oregoni méhfajra specializálódott, míg ugyan ennek a nünükének a Mojave-sivatagban élő populációja előbbi méhfaj egy itt élő, közeli rokonfaján, a Habropodapallida-n élősködik. A nünüke és a méhek közti kémiai verseny tetten érhető, ha (miként azt Leslie Saul-Gershenz és munkatársai tették) elemezzük az általuk termelt feromonkoktélok összetételét.
A hártyásszárnyúak szexferomonjai többnyire páratlan, 21-31 szénatomból álló, egyszeresen telítetlen szénhidrogének (a kettős kötés leggyakrabban a 9. szénatomnál található, de a különböző fajok feromonkoktéljaiban ettől eltérő pozícióba is kerülhet). A két Habropoda faj feromonjai között jellemző különbségeket találtak, és fajon belül természetesen a hímek és a nőstények ferromonprofilja is egészen másképp fest. Szépen tükrözi viszont a gazda-parazita kapcsolatot jellemző kémiai kódíró versenyt, hogy az oregoni M. franciscanus feromonjai között több olyan vegyület is található, ami a H. miserabilis nőstényére jellemző, míg a mojavei nünükék, noha biztosan ugyan abba a fajba tartoznak, mint az oregoniak (ezt anatómiai és genetikai elemzések is igazolják), feromonprofiljuk jelentősen eltérő. A mojavei populáció feromonkoktélja több olyan komponenst tartalmaz, ami a H. pallida-ra jellemző.
A H. miserabilis és a H. pallida hímjeinek és nőstényeinek valamint az oregoni és Mojave-sivatagi nünükelárvák feromon profilja. A vízszintes tengelyen a különböző szénatomszámú feromonok vannak feltüntetve, kék színnel jelölve azokat, amelyek legalább két csoportban ugyan úgy jelen vannak. a függőleges tengelyen relatív koncentráció látható (Leslie Saul-Gershenz és mtsai, 2017).
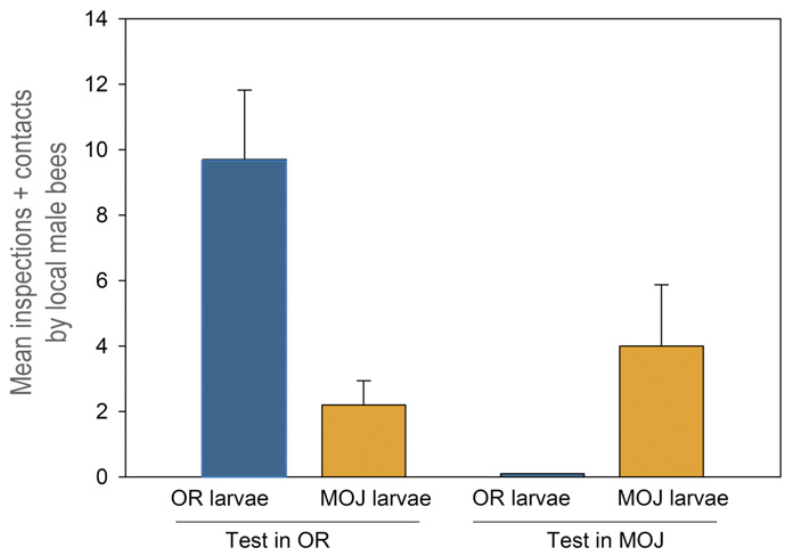
A feromonprofil vizsgálata mellett olyan kísérleteket is végetek a szerzők, amelyek a két nünükepopuláció eltérő gazdaspecificitását igazolták. Ezekből kiderült, hogy a mojavei nünükepopuláció valóban csak az ott élő Habropoda fajt vonzza magához, míg az oregonit pedig csak alig, vagy egyáltalán nem, és vice versa. Ezekről a kísérletekről olvasva az ember elgondolkozik rajta, hogy az eltérő gazdaspecificitás a két nünükepopuláció egyedei között (a jelentős földrajzi távolságokat nem számolva) mennyire járul hozzá a populációk szétválásához, vagy adott esetben magához a fajképződéshez…
A triungulinus lárvák eltérő gazdaspecificitását igazoló kisérletek eredményei (Leslie Saul-Gershenz és mtsai, 2017)
A jelen poszt forrásául szolgáló tanulmányok nem a tudomány legfrissebb termékei, és még csak azt se hiszem, hogy világmegváltó jelentőséggel bírnának. Amiért mégis beszámoltam róluk, annak csak az az oka, hogy idehaza épp „nünüke szezon” van. Bár a hazai Meloe fajok életciklusa kevéssé bizarr mint az M. franciscanus-é, ha a nyájas olvasó a hétvégén netán kirándulásra adná a fejét, és teszem azt egy kék nünükével (M. violaceus) találkozna gondoljon majd ezekre az érdekes amcsi rokonaira is 😉
Szép időt, és kellemes túrázást!
(A borítóképen egy kék nünüke látható – a szerző fotója)
Az állatvilágban (ha és amennyiben kétivarú fajokról beszélünk) a nemek kialakításáért felelős folyamatoknak (szexdetermináció) megdöbbentően széles spektrumával találjuk szembe magunkat. Csak a „magasabbrendű” magzatburkos gerincesekre szorítkozva eddig három egészen különböző logikájú rendszert írtak le: az emlősöknél és a madaraknál egyaránt az ivari kromoszómák játszák a fő szerepet, de míg előbbi esetben a hímek (XY), addig utóbbi esetben a nőstények (ZW) a különböző nemi kromoszómákat hordozó, ún. heterogametikus egyedek. A hüllők szerteágazó rokonságában szintén találkozhatunk kromoszomális szexdeterminációval, de a harmadik nagy kategóriát is a hüllők képviselői közt lelhetjük fel: sokuknál környezeti faktoroktól, pl. a fészekalj hőmérsékletétől függ a kifejlett példányok neme (nagyon sok hüllőfajt éppen ezért a klímaváltozás az ivararányok drasztikus eltolódása révén különösen érzékenyen érint). És persze előfordulhatnak előzőek furcsa kombinációi, mint a szakállas agámák esetén.
Adja magát a kérdés, hogy az egymással közelebbi, vagy távolabbi rokonságban álló magzatburkos csoportok milyen evolúciós utat járhattak be, ami végül az egyes csoportok eltérő ivarmeghatározó rendszereinek megjelenéséhez vezetett? Egy másik érdekes kérdés, hogy a sokféle szexdeterminációs rendszer közül melyik hasonlíthat legjobban a magzatburkosok utolsó közös ősében működő rendszerhez. A kérdés megvizsgálására a modern genetikai eszközök és új tudományágként az összehasonlító genomika kiváló lehetőségeket kínál.
Vegyük rögtön (nem minden nárcizmustól mentesen) a saját példánkat! Ha kíváncsiak vagyunk rá, hogy honnan származik a Homo sapiens ivarmeghatározása, akkor először körbe kell nézni a méhlepényes emlősök (Placentalia) között, majd vetni kell egy pillantást az erszényesek (Marsupialia) szexkromoszómáira, végül meg kell nézni az emlősök osztályának legkorábban (kb. 160-180 millió éve) leágazott rendjét, a kacsacsőrű emlőst (Ornithorhynchus anatinus) és a 4 recens hangyászsünfajt magába foglaló Monotremata csoportot.
Ahogy az emlősök családfájának ágain ugrálva távolodunk saját fajunktól, úgy végigkövethetjük az X és az Y kromoszóma változásait is, de ha leérünk a fa legalsó ágához, ott nagy meglepetések fogadnak. A Monotremata csoportnak ugyanis nem pusztán 1, hanem rögtön 5 pár szexkromoszómája van. Ez az öt pár kromoszóma ráadásul egy nagyon különleges multikromoszómás komplexet alkot a sejtosztódások során, megelőzendő a homológok véletlenszerű kiosztását a leánysejtek között. Különösen fontos ez az ivarsejteket képző számfelező osztódás (meiózis) során, hisz egy ivarsejtbe (gaméta) vagy csak X, vagy csak Y kromoszómák kerülhetnek.
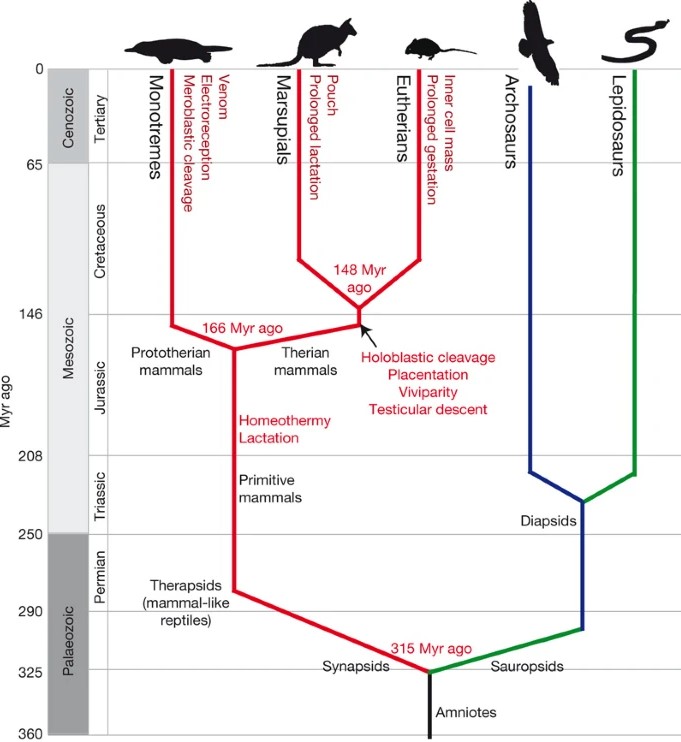
A Monotremata, Marsupialia és a méhlepényeseket is magába foglaló Eutheria csoportok viszonyait mutató, néhány fontos evolúciós újítást is feltüntető törzsfa (Warren és mtsai. 2008)
A Monotremata szexkromoszómák (miként más állatoknál is) tartalmaznak nem rekombinálódó, a nemek kialakítását szabályozó régiókat (SDR – sex determination region), illetve olyan régiókat, amelyek lehetővé teszik a kromoszómapárok egymásra találását – ezek a régiók ugyanúgy működnek, mint a nem-kromoszómák, azaz automszómák (nevük is ennen ered: pszeudoautoszómális régiók – PAR). A trükk, ami a különleges multikromoszómás komplex kialakulását eredményezi abban áll, hogy nem csak a kromoszómapárokon, de a sorban egymást követő X és Y kromoszómákon is találhatók egymáshoz illeszkedő PAR-ok. Így tehát a kacsacsőrű emlős X1-es kromoszómája nőstény egyedekben kapcsolódhat egy másik X1-hez és X2-höz is, míg hímekben az egyik X1 helyett Y1 áll. Az Y1 X2-höz, az X2 Y2-höz és X3-hoz, X3 Y3-hoz és X4-hez, ami Y4-hez és X5-höz, ami végül Y5-höz illeszkedik meióziskor.
További érdekesség, hogy a szekvenciaelemzések az így kialakuló kromoszómaláncok két végén is feltártak egy-egy, mostanra már erősen mutálódott PAR elemet, amelyek a mai Monotrematák őseinél elvileg lehetővé tehették, hogy a láncszerű multikromoszómás komplex gyűrűvé záródjon. Ha ez a hipotetikus őskromoszóma-komplex valóban képes volt a saját farkába harapni, az már csak azért is izgalmas, mert eddig az egész élővilágban csak egy növényél, pontosabban egy ligetszépe fajnál figyeltek meg gyűrűbe záródó meiotikus kromoszómaláncot. Ritka kuriózum hát az ilyen!
A kacsacsőrű emlős és a hangyászsün genomok feltérképezésének korai szakaszában, Rens és munkatársai közöltek egy izgalmas összehasonlító tanulmányt, amelyben akkor főleg in situ hibridizációs (FISH) módszerek segítségével vetették össze egymással a kacsacsőrű emlős és a hangyászsünök, valamint a méhlepényesek és a madarak ivari kromoszómáit. Ami már az elején feltűnő volt, hogy a Monotremata fajok szexkromoszómái néhány apróbb különbséget leszámítva erősen konzerváltak. Az egy már korábban megfigyelt különbség, hogy a hangyászsünök Y5-ös kromoszómája hiányzik. Ez a kromoszóma a kacsacsőrű emlősnél is meglehetősen apró, így korábban elfogadható magyarázatnak számított, hogy a hangyászsünök egyszerűen elveszítették azt valamikor az evolúciójuk során. Rensék kutatásai annyival árnyalták ezt a képet, hogy a hangyászsün Y3-as kromoszómáján találtak olyan régiókat, amelyek az elveszettnek hitt Y5-nek felelnek meg. Az újabb értelmezés szerint tehát ez a kromoszóma nem eltűnt, csak integrálódott a hármas sorszámú Y kromoszómába, hasonlóan ahhoz, amikor két kártyapaklit egybekeverünk.
A rövidcsőrű hangyászsün (Tachyglossus aculeatus) és szexkromoszómái, amint láncszerű komplexet alkotnak (Rens és mtsai. 2007).
Visszatérve eredeti kérdésünkhöz, hogy honnét származik a méhlepényesek, illetve az ember X és Y kromoszómája, egy dolgot már biztosan elmondhatunk: nem a legelső emlősöktől örököltük őket. A Monotremata szexkromoszómák ugyanis nem mutatnak semmilyen homológiát a Placentalia szexkromoszómákkal. A mi magányos és aprócska Y kromoszómánkon található SRY lókusz pedig, ami a méhlepényeseknél kulcsszerepet tölt be a herék fejlődésének kezdetén, s így végső soron a hím nemi jellegek megjelenésének forrása, mind a kacsacsőrű emlős, mind pedig a hangyászsünök genomjából hiányzik.
A méhlepényesek X kromoszómájának hosszú karján található egy, a dóziskompenzációban fontos lókusz, az XIST. Ennek a szakasznak a megfelelőjét kimutatták ugyan a kacsacsőrű emlős genomjában, de nem az egyik ivari kromoszómán, hanem a 6-os autoszómán, arra pedig egyelőre semmi nem utal, hogy ez a lókusz bármiféle szerepet is betöltene a kacsacsőrű emlősök szexdeterminációjában. Jelenleg úgy tűnik tehát, hogy a Monotremata és a többi emlős szexdeterminációs rendszere egymástól függetlenül jelent meg a két csoport elválását követően, és semmiféle közös eredettel nem bír.
Nagyfokú homológia mutatható ki azonban a kacsacsőrű emlős X5-ös, a hangyászsünök X4-es szexkromoszómája, valamint a madarak Z kromoszómája között. A madarak szexdeterminációjában fontos szerepet játszó, dózisfüggő transzkripciós faktorok, a DMRT-1, -2 és -3 mind megtalálható a kacsacsőrű emlős X5-ös szexkromoszómáján, de találtak olyan Z kromoszómára térképezett géneket is, amelyek ortológjai a kacsacsőrű emlős X2 és X3 ivari kromoszómáján helyezkednek el. Ugyanezek a Monotremata kromoszómarégiók nagyfokú egyezést mutattak a humán 9-es autoszómával is, ami többé-kevésbé megfeleltethető a madarak Z kromoszómájának, illetve a humán 5-ös és 18-as kromoszómáknak azon részleteivel, amelyek a madarak 2-es autoszómájával hibridizáltathatók.
Azt jelentené ez, hogy a Monotremata és a madár szexdetermináció (az XY és a ZW rendszerek eltérő logikája ellenére) azonos gyökerekre vezetnek vissza? A hüllőket és madarakat magába foglaló Sauropsida, valamint az összes mai emlőst, és egy seregnyi emlősszerű őshüllőt tartalmazó Synapsida csoportok több mint 300 millió éve, a késő Karbonban válhattak el egymástól. Ha tehát elfogadjuk az iménti felvetést, azzal azt is feltételezzük, hogy a legkorábbi magzatburkosok nemmeghatározása szempontjából a DMRT gének fontosak lehettek.
Árnyalja a képet egy idén januárban a Nature hasábjain megjelent cikk, amelynek szerzői már elég részletes genomi információk birtokában kísérelnek meg rendet tenni az emlős kromoszómák háza táján. A kísérlet nem volt sikeres, hisz a rendcsinálás közben a kupleráj, amelybe a fenti sorok olvasása közben betekintést nyerhettünk, csak sokszorosára duzzadt.
A fali gyík, a házityúk, a kacsacsőrű emlős, a tasmán ördög, egy oposszum faj és az ember genomjának konzervált kromoszómális régióit és e régiók egymáshoz viszonyított pozícióját elemezték, s megállapították, hogy e régiók igencsak össze lettek keverve az elröppent év-százmilliók alatt. A szerzők ötszáznál is több független kromoszómaátrendeződési eseményt azonosítottak (transzlokációk, kromoszómafúziók és fragmentálódások).
Most képzeljük el, hogy valaki egyszer az aztalon felejtett néhány pakli kártyát, amiket minden arra járó ember keverget egy kicsit. Nekünk pedig egyszer csak a mániánkká válik, hogy lépésről-lépésre megértsük a paklik összekeveredésének történetét. Szerencsénkre a kártyaasztal a rajta lévő paklikkal együtt, valami rejtélyes módon párszor lemásolta saját magát, így az egyes másolatokból kinyerhetünk némi információt arra vonatkozóan, hogy a másolatkészítés pillanatában hogyan néztek ki az „ősasztal” „őskártyapaklijai”. De jaj! Valaki aljas módon tovább keverte az összes másolat-kártyapaklit is! Ön hogy állna neki az eredeti kártyapaklik rekonstrukciójának?
A rekonstruált emlős-őskromoszómák (MAC), és a kromoszómaátrendeződési események az egyes leszármazás ágakon, színkódolva. Az ágakon a beazonosított események száma, az elágazási pontoknál pedig az ott jelölt közös ős diploid kromoszómaszerelvénye látható (Zhou és mtsai. 2021).
A megoldás kulcsát az asztalmásolatok jelentik. Ha ezeket összehasonlítva az egyes paklik kártyasorrendjében hasonlóságot találunk, akkor az a másolatkészítés előtt alakult ki, ha pedig különbségeket, akkor azok olyan kevergetésből származnak, amelyek a másolatkészítést követően történtek. A kártyás allegóriát feloldva: a kártyalapok kromoszómarégiókat, a paklik kromoszómákat, az asztalok pedig fajokat jelölnek, amelyek összehasonlítása révén nyílik lehetőségünk a keverés történetének és végső soron az „őskártyapaklik” rekonstrukciójára. Zhou és munkatársai elvégezték hát ezt a heroikus rekonstrukciót, és egyebek mellett megállapították, hogy az emlősök közös ősének valószínűleg 30 pár kromoszómája volt. A szerzők azon túl, hogy mind a 30 őskromoszómáról (Mammalian Ancestral Chromosome – MAC) leírták a konzervált régiók relatív sorrendjét, fontos megállapításokat tettek a szexkromoszómákkal kapcsolatban is.
Úgy tűnik, hogy a Monotremata 5 pár ivari kromoszómájának mindegyike több, különböző kártyapakli lapjaiból tevődött össze. Az a hasonlóság tehát, amit anno Rens és szerzőtársai a madarak és a Monotremata szexkromoszómák között megfigyeltek szinte biztosan nem arra vezethető vissza, hogy a madár Z és a kacsacsőrű emlős X5 kromoszómája egy közös őskromoszóma két távoli leszármazottja lenne.
Az új tanulmányok fényében az a fentebb megfogalmazott elmélet is megingott kissé, hogy DMRT gének ivari determinációban betöltött szerepe egy ősi magzatburkos vonás lenne. Hiszen egy ilyen viharos sorsú kártyaasztalnál nem elképzelhetetlen az sem, hogy két független kiosztásban kap egy gén hasonló szerepet, s kerül mondjuk szexkromoszómára. Tartsuk szem előtt, hogy a madarak és az emlősök utolsó közös őse több mint 300 millió éve élt a Földön, és hogy ismerünk olyan állatokat is, ahol egy fajon belül kettő, vagy több tök különböző szexdeterminációs rendszer is működik!
Na és mit tudunk meg végre a saját ivari kromoszómáinkról?
Korábban említettük az Y kromoszómánkon található SRY lókuszt, amely a Theria (tehát erszényes és méhlepényes) közös ősben már jelen volt és a maihoz hasonló szerepet tölthetett be, de a Monotremataban még nyomát sem látni. Az X kromoszómánk szintén a Monotremata-Theria elvállást követően jelenhetett meg, de a méhlepényeseknél fontos dóziskompenzációs gén, az XIST csak a méhlepényes-erszényes elvállást követően bukkant fel az X kromoszóma történetében. Fontos különbség még a kenguruk és az ember X kromoszómája között, hogy a méhlepényesek X kromoszómája az ősidőkben fuzionált egy ősi Theria autoszómával, míg az erszényeseknél ilyen fúziós esemény nem történt.
Szövevényes történetek játszódhattak a kártyaasztalok körül, és egyenlőre csak a legfontosabb események vázlatát látjuk. Ugyanakkor nem tudunk semmit azokról a játékosokról, akiknek a lapjárása olyan szerencsétlenül alakult, hogy kihajították őket az evolúció kaszinójából, és azok se buktak még le, akik cinkelt kártyával játszottak. Egyszer majd talán ezekre a történetekre is fény derül.
__________
Nicole Valenzuela és mtsai. (2019) Extreme thermal fluctuations from climate change unexpectedly accelerate demographic collapse of vertebrates with temperature-dependent sex determination” Scientific Reports
Craig A Smith és mtsai. (2009) The avian Z-linked gene DMRT1 is required for male sex determination in the chicken Nature
Hieronim Golczyk és mtsai. (2014) Translocations of Chromosome End-Segments and Facultative Heterochromatin Promote Meiotic Ring Formation in Evening Primroses. Plant Cell
Willem Rens és mtsai. (2007) The multiple sex chromosomes of platypus and echidna are not completely identical and several share homology with the avian Z. Genome Biology
Yang Zhou és mtsai. (2021) Platypus and echidna genomes reveal mammalian biology and evolution. Nature
A véletlenszámok generálása mindig is a hétköznapok szerves részét képezte. Már az ókori Egyiptomból és Mezopotámiából (i.e. 3000 környéke) is maradtak fenn olyan népszerű társasok, ahol a fej-vagy-írás a játék központi mechanizmusa. Sokkal később már volt életművész, aki a fizikai Nobel-díjat a makao nevű kártyajátékon nyerte el Noah Bertinus professzortól. Az én gimnazista éveimnek pedig legizgalmasabb pillanatai közé sorolható, mikor d10-esekkel (10 oldalú dobókocka) egész orkhordákat tettünk el láb alól, ha a sors nekünk kedvezett. Akkor még ciki volt szerepjátékozni, aztán jött a Trónok Harca és a Stranger Things…
Manapság azonban a kocka és érme már nem elég hatékony. Számítógépes szimulációk során például másodpercenként több ezer random számra is szükség lehet. De talán ennél is fontosabb ipar napjainkban a kriptográfia (meg vannak véletlensevendéglátóegység kaszinók). Legyen szó banki átutalásról vagy bármilyen kommunikációról a digitális térben, a bizalmas információk titkosítása hatalmas nagyságrendben igényel véletlenszámokat.
Pszeudo és valódi random számok
Léteznek úgynevezett pszeudo (nem valódi) random számok. Könnyen lehet ilyeneket generálni egyszerűbb matematikai szabályokkal is, amik egy kezdeti értékből kiindulva randomnak tűnő számsorozatokat eredményeznek. A legtöbb területen (modellezés, számítógépes játékok) ezek is megállják a helyük. Azonban még a komolyabb pszeudorandom generátorok is rejtenek magukban hátrányokat. Legnagyobb veszélyük, hogy determinisztikusak, azaz ha valaki ismeri az algoritmust és generátor belső állapotát, akkor meg tudja jósolni, hogy az egyes lépésekben milyen számok keletkeznek. Virtuális lottósorsolásokon hasonló manőverrel már nyert valaki körülbelül 7,5 milliárd forintnak megfelelő dollárt (majd 25 év letöltendőt), illetve az Amerikai Nemzetbiztonsági Ügynökséggel (NSA) kapcsolatban is felmerült, hogy egyes titkosításra használt számgenerátorokba ilyen kiskapukat épít.
Pont ezért olyan izgalmas piac a valódi random számok területe. Ezek a számgenerátorok szoftveres és algoritmikus megoldások helyett ténylegesen véletlenszerű fizikai folyamatok valós idejű mérésére építkeznek. Az eképpen keletkezett adatok teljesen jósolhatatlanok, így biztonságosabbak is kriptográfiai szempontból. Ilyen véletlen forrása lehet, a teljesség igénye nélkül, a radioaktív bomlás, atmoszférikus rádiózajok, áramkörökben fellépő fluktuációk és egyéb kvantummechanikai események (ezek közül a kedvencem a kaotikus félvezető lézer, ami úgy hangzik, mint a hatodik Halálcsillagra tervezett szuperfegyver).
D&D: DNS és Digitalizációja
Egy ilyen, valódi véletlenszám-generátorokkal kapcsolatos felvetés, hogy kémiai folyamatokat molekuláris szinten figyeljünk meg, és ezt használjuk ki valamilyen módon. Itt jön a képbe a DNS, mint egy lehetséges rendszer, melyet mostanság sikerült megvalósítani Meisernek és munkatársainak.
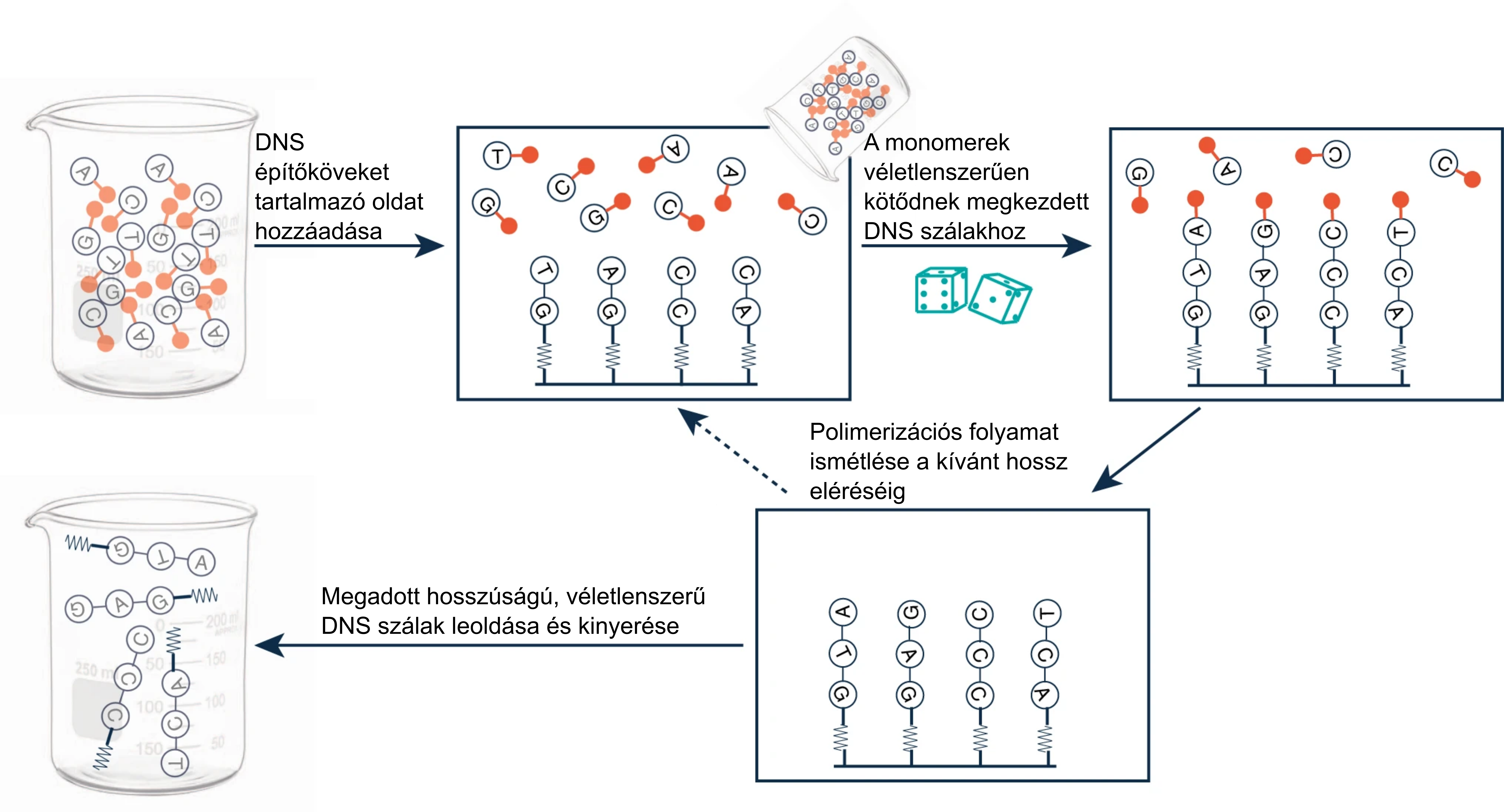
Hogy zajlik ez gyakorlatban? Vegyük e nukleinsav építőköveit, a négyféle dNTP-t (leánykori nevükön dezoxiribonukleotid-trifoszfátokat): dATP, dTTP, dCTP és dGTP. Ezek elegyéből indulunk ki egy véletlenszerű DNS szintézise során. Nem egy meglévő DNS-t másolunk; minden egyes polimerizációs lépés olyan, mintha dobnánk egy négyoldalú dobókockával: először mondjuk egy G-t, aztán A-t, T-t, újra T-t satöbbi. Így jutunk egy random DNS-szálhoz (1. ábra). Sőt, nem is kell nekünk szenvedni a laborban! Random DNS-ek már kereskedelmi forgalomban is kaphatóak, ha valaki szeretne egyet otthonra. Pont olyan izgalmas a tartásuk, mint egy cserép petúniának.
1. ábra: Random DNS-ek szintézise (forrás:Meiser és mtsai., 2020)
Ami ezek után még hátravan, az a generált random DNS-ek beolvasása (szekvenálás) és random számokká alakítása. A szekvenálás manapság elterjedt újgenerációs technológiákkal könnyen és gyorsan megoldható. A DNS-ek random számmá való átdolgozása sem kihívás. A digitális világ amúgy is bináris értékekkel operál. Rendeljünk mindegyik bázishoz nullát vagy egyet a következőképpen: A→0, C→0, T→1, G→1. Így lesz mondjuk a GATTACA szekvenciából 1011000, mint random bitek sorozata.
Az olvasóban felmerülhet, hogy megduplázhatnánk a hatékonyságot, ha kettesével osztanánk ki a biteket, pl.: A→00, C→01, T→10, G→11. Ennek hiánya a jelenlegi technológia egyik gyermekbetegségének, pontosabban annak megoldásának tudható be. A random DNS szintézis ugyanis nem teljesen random, G és T sokkal gyakrabban épül be a jelenlegi eljárások során. Ez azt is jelenti, hogy bizonyos véletlenszámok nagyobb valószínűséggel ütnék fel a fejüket. A probléma más számgenerátorok esetében is ismert, megoldására létezik is algoritmus, melyet von Neumann korrektor-nak hívnak (Neumann János után). Ez a korrekció azonban nem működne, ha kettesével generálnánk a biteket.
Ami ennél is fájdalmasabb, hogy a korrektor is további veszteségekkel dolgozik, az eredeti nemannyiravéletlen bittengerből így csak negyedannyi igazánvalódi random számot lehet kitermelni. Összességében tehát nyolcszoros veszteségről beszélhetünk. Mégis, az ilyen csiszolatlanságai ellenére is elfogadhatóan teljesít a DNS ha más generátorokkal vetjük össze (1. táblázat). Különösen igaz ez akkor, ha azt az ismertebb valódi véletlenszám technológiák teljesítményéhez mérjük.
1. táblázat: DNS és más véletlenszám-generátorok teljesítménye, Meiser és mtsai. (2020) nyomán, módosítva
A DNS-alapú rendszer komoly előnye a könnyű hordozhatósága: egy 1ml-nyi DNS körülbelül 1019 bit információt tud raktározni (ez merevlemeznek sem utolsó, de a mi esetünkben ez mind értékes random bit lehetne). Szekvenátorokból is vannak már egész kompakt típusok. A rendszer jelenlegi korlátját és fő költségét is ez utóbbi, a szekvenálás jelenti, ami viszont továbbra is egy fejlődő terület. Javulásra tehát még számíthatunk a későbbiekben, s így semmi jövőbeli akadálya nincs annak, hogy a holnap társasjátékait már sufnituning dNTP random számokkal játszhassuk.
Reidler, I., Aviad, Y., Rosenbluh, M. & Kanter, I. Ultrahigh-speed random number generation based on a chaotic semiconductor laser. Phys. Rev. Lett.103, 1–4 (2009). https://doi.org/10.1103/PhysRevLett.103.024102
Zhirnov, V., Zadegan, R., Sandhu, G. et al. Nucleic acid memory. Nature Mater15, 366–370 (2016). https://doi.org/10.1038/nmat4594
Az organoidok, vagyis miniatűr, in vitro (lásd még “Petri csészében”) létrehozott szövetdarabok kutatása az utóbbi években mindenképpen a leggyorsabban fejlődő biológiai tudományterületet jelenti és manapság már tényleg (hogy elpuffantsuk az idekívánkozó nagy közhelyet) nem múlik el hét 1-2 fajsúlyosabb organoidos publikáció nélkül.
Madártávlatból nézve, két nagy csoport különíthető el organoidok esetében: azok, amelyek nagyon széles differenciációs potenciállal rendelkező “pluripotens” (akár klasszikus, embrionális, akár újgenerációs indukált) őssejtekből jönnek létre és azok, amelyek szűkebb potenciállal rendelkező, “multipotens” őssejtekből alakulnak ki. Utóbbiak döntően a felnőtt, kifejlett egyedek egyes szöveteiben jelen levő őssejtek felhasználásával létrehozott organoidok és ennek a területnek az egyik legfontosabb úttörője a pár éve Breakthrough Prize-al is kitüntetett Hans Celvers.
Clevers csoportja a holland Hubrecht Institute-ban még a 2010-es évek elején ért el komoly áttöréseket a bél-organoidok létrehozásával és kutatásával, azóta pedig, talán csak minimális túlzással, nincs olyan hámszövettel (epitéliummal) borított szerv, amelyből ne készítettek volna organoidokat. Az utóbbi években különböző mirigyekre kezdtek koncentrálni, ehhez kapcsolódott egy tavalyi publikációjuk, ahol például méreg-termelésre képes kígyó méregmirigy-organoidokat csináltak. Most pedig, a jól bejáratott metodológiájukkal a könnymirigyet vették górcső alá.
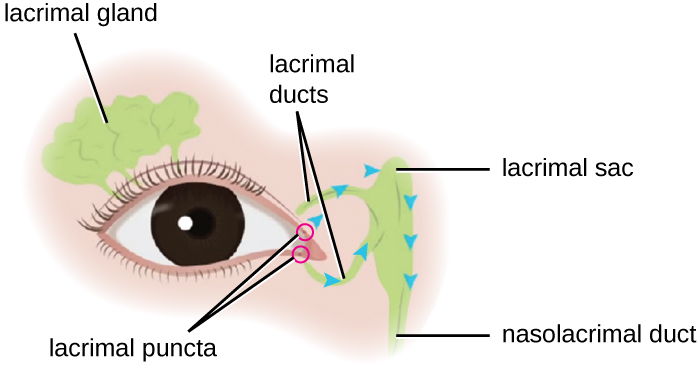
Az emberi könnytermelésben szerepet játszó mirigy és csatorna. (Forrás: Wikimedia Commons)
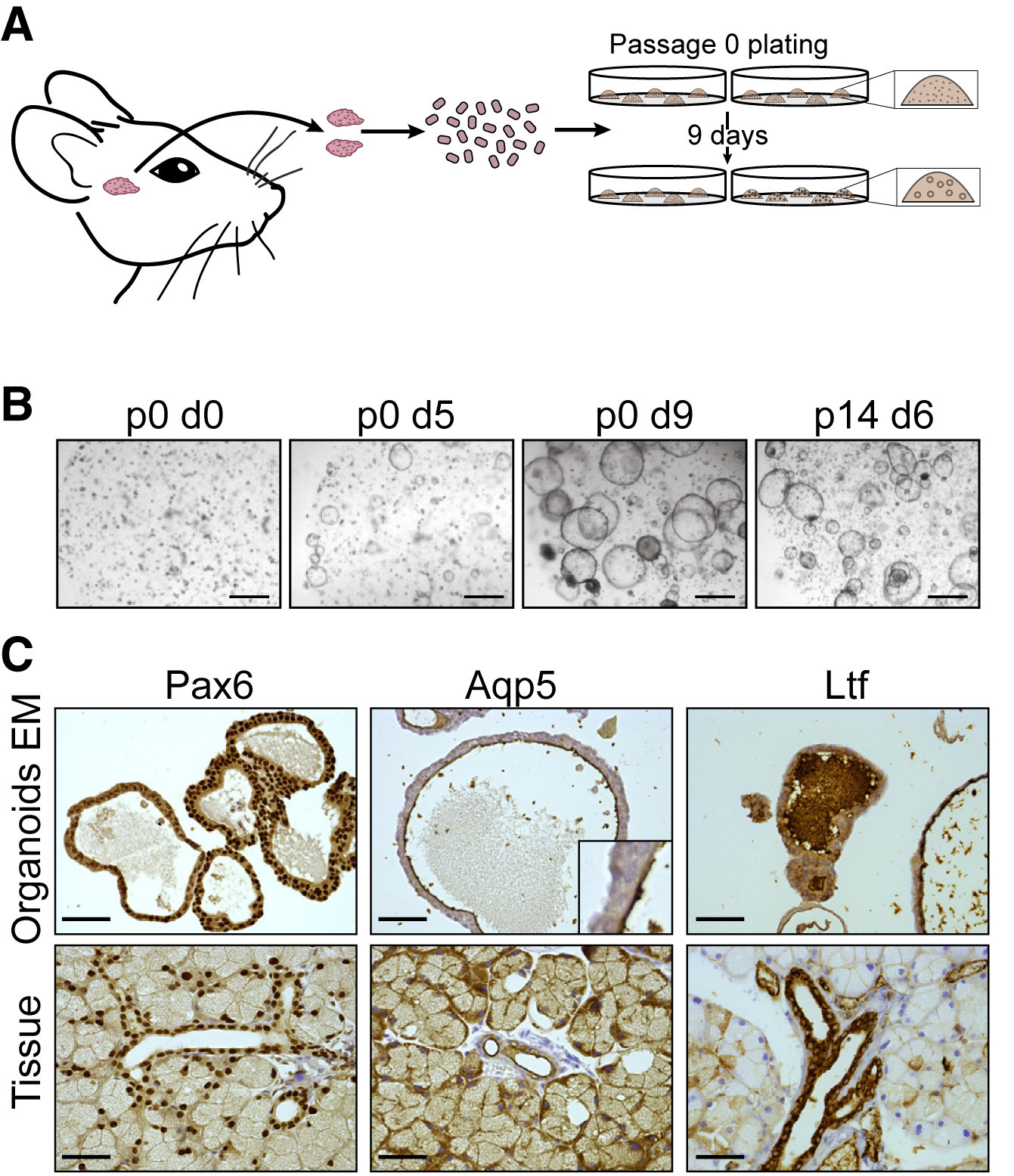
Az ilyen organoid-készítős tanulmányoknak a kezdeti lépései mindig nagyon hasonlóak: a vizsgált szervet szétszedik sejtekre, amelyeket utána egy in vitro közegben próbálnak meg tenyészteni. Ha a sejtek között vannak őssejtek is, akkor a megfelelő közeg kikísérletezése után előbb-utóbb megjelennek a labda alakú organoidok. Most is ez történt, előbb emberi, majd egér könnymirigyből kiindulva hozták létre ezeket az organoidokat (alább csak az egér esetet mutatom), amelyek kb. 10 nap alatt ∼200 μm átmérőjű gömbökké nőtték ki magukat és a szövettenyészetek fenntartására klasszikusan használt “passzálások” (újabb tenyészet indítása a sejtekből) után is megmaradt ez a jelleg (az ábra B paneljén a “pX” azt jelenti, hogy X db. passzálás után; a “0” azt jelenti, hogy a frissen létrehozott szövetet jellemzik.) Az organoidok több olyan fehérjét kifejeztek, ami a könnymirigyre is jellemző, sőt a belsejükben, hasonlóan a valódi könnymirigyek belsejéhez az antimikrobiális hatású Lactotransferrin (Ltf) is megjelenik.
Ha pedig van egy olyan organoid-rendszerünk, amely gyakorlatilag szinte korlátlan mennyiségben (viszonylag olcsón) tudja biztosítani egy valódi szerv miniatűr vázát, hirtelen a lehetőségek elég széles tárháza nyílik meg. Nagyon részletesen lehet vizsgálni a szerv kialakulásának folyamatát, az egyes gének szerepét (ezáltal genetikai-eredetű betegségek kialakulását), vagy azt is, hogy az organoidot visszaültetve, mennyire lesz működőképes (ez potenciális terápia lehet a jövőben pl. Sjögren-szindróma típusú autoimmun betegségekre). Én csak egy aspektust emelnék ki ebből a tanulmányból, mégpedig azt, hogy az organoidok miképp tehetik lehetővé a könnyezést befolyásoló szerek gyors keresését.
Könnymirigy-organoid “duzzadási vizsgálatok menete (A), konkrét anyagokkal való tesztelés (B), ennek a kvantifikációja (C) és a noradrenalin (NorEpi) feltételezett szerepe a könnyképzésben. (Forrás: Cell Stem Cell)
A munka ezen részéhez már emberi könnymirigyből származó organoidokat használtak és lényegében arról van szó, hogy különböző anyagokat tettek az organoidok közegéhez. Normális könnymirigynél a könny az elvezetődik, de az organoid egy zárt rendszer, így a megtermelődő, az organoid belsejébe ürülő könny lufiként “felfújja” az organoidokat. Ezt a méretváltozást pedig elég könnyen lehet automatizálva is detektálni, s ahogy az a fenti ábrán is látható, elég gyorsan lehet szűrni azokat az anyagokat, amelyek könnyezést tudnak indukálni.
Miért lehet ez hasonló, miért akarnák elérni, hogy valaki könnyezzen (és miért nem hámoztatunk vele hagymát)? A helyzet az, hogy bár egy könnymirigy anatómiailag elég egyszerű rendszer, még nem értjük teljesen, miképp működik. Ha pedig olyan anyagokkal, amelyeket korábban már pontosan jellemeztek (tudjuk pl. milyen molekulákhoz kapcsolódnak és befolyásolnak) sikerül befolyásolnunk, közelebb kerülhetünk ahhoz, hogy megértsük, a valóságban hogyan történhet a könnymirigy működésének szabályozása.
Korving J, Trani Bustos M, Gehart H, Begthel H, Bar-Ephraim YE, et al. (2021) Exploring the human lacrimal gland using organoids and single-cell sequencing. Cell Stem Cell doi: 10.1016/j.stem.2021.02.024.
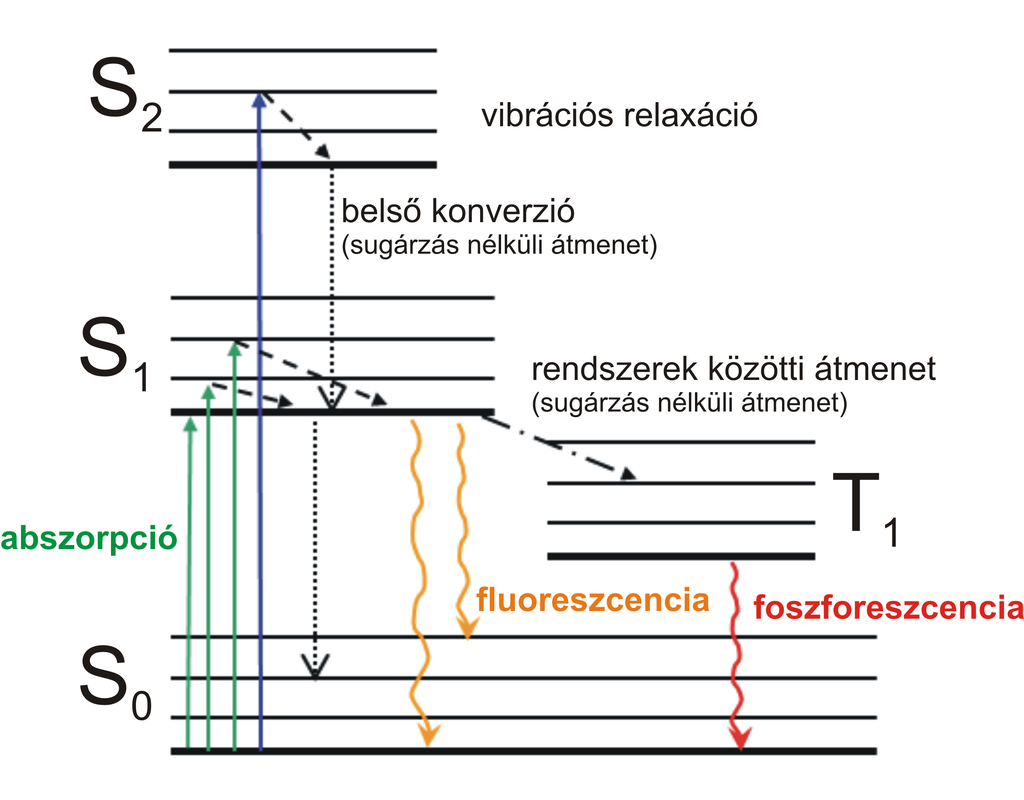
Az élővilág megszámlálhatatlan csodái közül talán az egyik legmisztikusabb a biolumineszcencia és a fluoreszcencia. E két jelenség alapvetően eltérő okokra vezethető vissza, és hogy itt együtt említem őket, annak csupán az az oka, hogy mindkét esetben valamiféle fényjelenséggel van dolgunk. A fény forrása előbbi esetben mindig egy kémiai reakció, aminek végbemenetelét különböző enzimek – biológiai katalizátorok – segít(het)ik, míg utóbbi esetben kémiai változás nem történik, a fény eredeti forrása pedig kevésbé hangsúlyos. A fluoreszcencia lényege abban áll, hogy meghatározott hullámhossztartományba eső fotonok kölcsönhatásba lépnek egyes molekulák gerjeszthető elektronjaival (abszorpció). Az elektronok a kölcsönhatást követően magasabb energiaállapotba kerülnek, majd pár röpke nanoszekundum elteltével apró rezgések és mozgások révén veszíteni kezdenek energiájukból (belső konverzió), majd visszazuhannak a gerjesztés előtti alapállapotba, miközben egy foton lép ki a molekulából (emisszió). A kibocsájtott foton energiája, minthogy a gerjesztett állapotú elektron energiája egy részét mindenféle zizegésekre és mocorgásokra pazarolja, mindig kisebb, mint az elnyelt fotoné. A fluoreszkáló felületekről származó fény tehát mindig alacsonyabb energiájú és nagyobb hullámhosszú, mint a gerjesztéshez használt fény; más szavakkal élve: a szivárvány pirosabbik oldala felé tolódik el.
Az elektronok energiaszintjének változásait és a fluoreszencia alapjait sematikusan ábrázoló Jablonski-féle diagram (kép forrása: Wikipédia)
A biolumineszcenciára kiváló példát szolgáltatnak a szentjánosbogarak, számtalan mélytengeri ketyere, köztük egy Swima bombiviridis nevű gyűrűsféreg, akiről korábban itt írtunk, valamint korallok, medúzák, algák és baktériumok egész sora. Az is előfordul, hogy egy állat egyszerre villantja fel a biolumineszcencia és a fluoreszcencia területén szerzett jártasságát is, mint a zöld fluoreszcens fehérjéről (GFP) méltán világhírű Aequorea victoria nevű medúzafaj.
Fluoreszkáló állatoknak is komoly sokaságát lehetne felsorolni. Az ízeltlábúak testét borító kitines kutikula, vagy egyes kétéltűek (pl. a Hypsiboas punctatus nevű Dél-amerikai levelibéka faj) bőrében termelődő mirigyváladék egyaránt bírhat fluoreszcens sajátságokkal. A szárazföldi gerincesek körében nem gondolnánk, hogy a fluoreszcencia gyakori jelenség lenne, pedig elég egy pörgősebb party-ba ellátogatnunk, hogy erről első kézből szerezzünk tapasztalatokat: fogaink (és kevésbe feltűnő módon csontjaink is) kékesen világítanak az UV fényben (a csontszövet emissziós maximuma a 430 nm körüli hullámhossztartományban van, de remélhetőleg a party sose durvul el annyira, hogy ezt módunkba álljon saját magunkon megfigyelni).
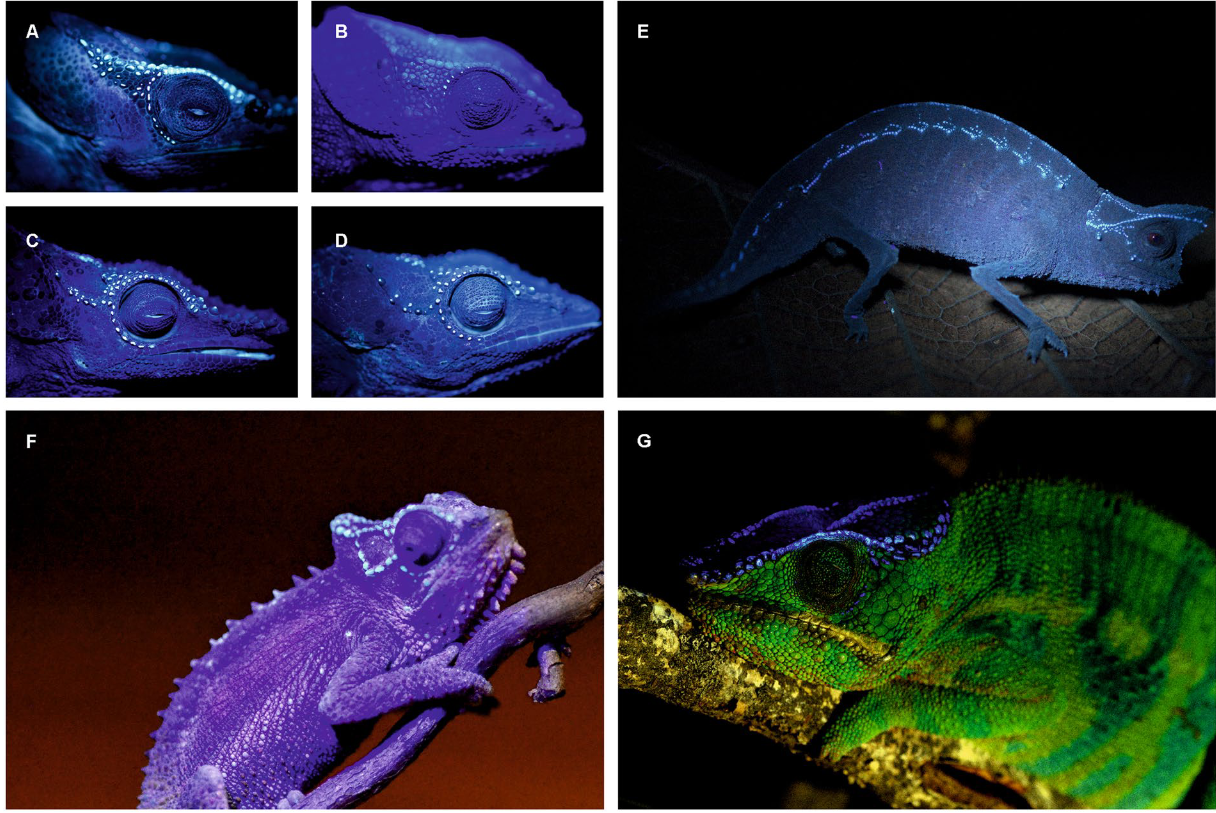
A csontszövet immanens fluoreszcenciáját több, elsősorban éjszaka aktív állatfaj is felhasználja. Példa erre a Délkelet-Brazíliában honos nyergesbékafélék (Brachycephalus) nemzetsége, amelyek két fajánál is leírták, hogy a fej és a hát területén a bőrben, szinte közvetlenül a felhám alatt erőteljes csontosodási folyamatok mennek végbe. Minthogy ezek a bőr eredetű, ún. dermális csontok a testfelszínhez rendkívül közel helyezkednek el, az UV fény képes a testfalon át elérni azokat, s a kibocsájtott kék fény is gond nélkül lép ki a csontlemezeket fedő vékony szövetek borításán keresztül. Hasonló módon nyűgözik le egymást a Calumma és Bradypodion nemzetségbe tartozó kaméleonok is. Ezeknél a hüllőknél a koponya apró csontkinövései kerülnek a testfelszín közvetlen közelébe, a fajra jellemző mintázatban.
Néhány kaméleonfaj, fluoreszkáló csontkinövésekkel: A) Calumma crypticum, hím; B) C. crypticum, nőstény; C) C. cucullatum, hím; D) C. cucullatum, nőstény és E) Brookesia superciliaris, hím példányok 365 nm-es UV-fénnyel megvilágítva, valamint F) Bradypodion transvaalense és (G) Furcifer pardalis, hím példányok (gyenge, látható fényben, 365 nm-es UV fénnyel megvilágítva) (Prötzel és mtsai, 2018)
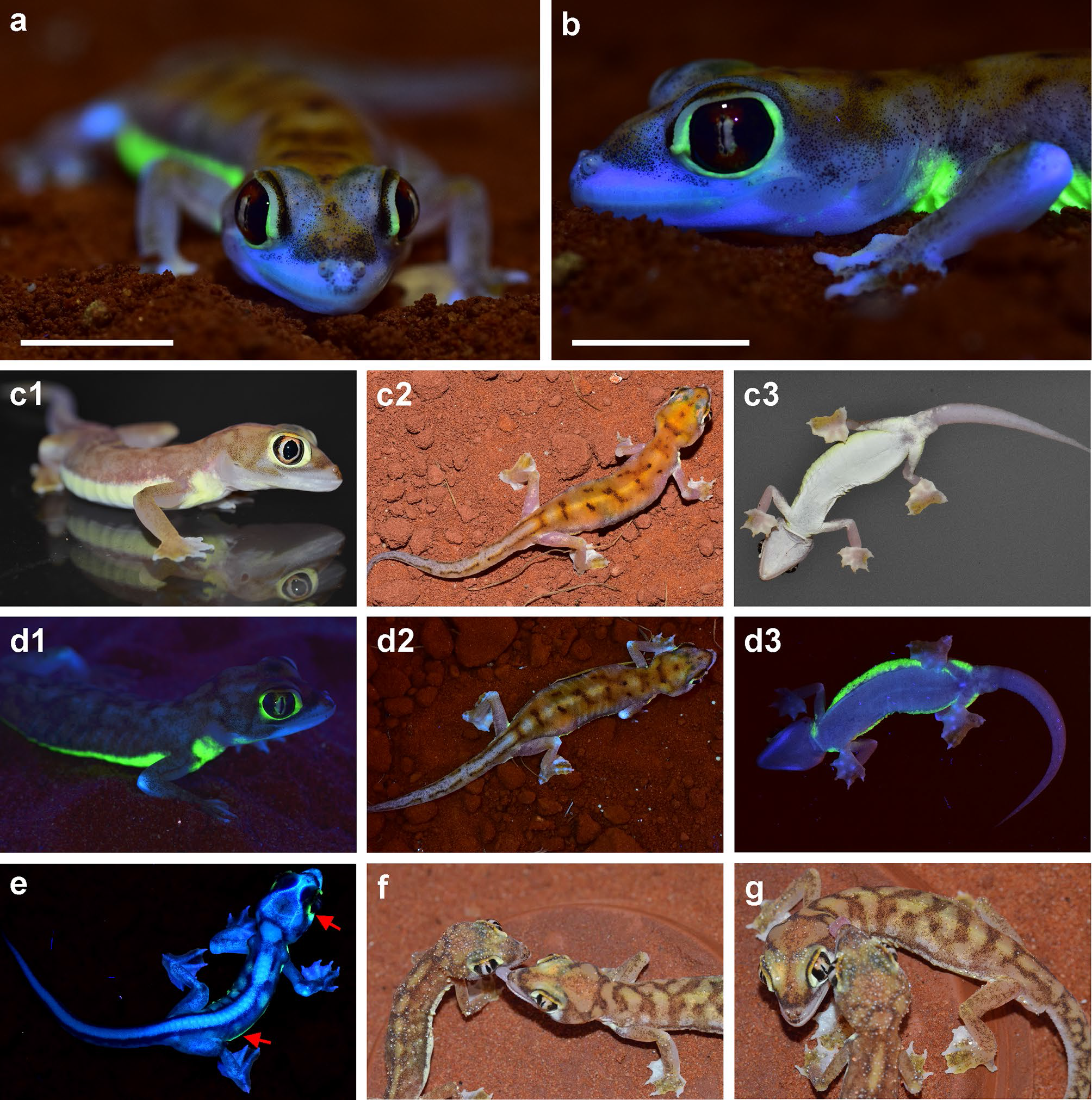
Jelen poszt apropóját egy idén januárban megjelent cikk adja, ugyan azoknak a szerzőknek a tollából, akik a fenti képen látható kaméleonokat is zaklatták UV-lámpáikkal. A szerzők megfigyelték, hogy a Namib-sivatagban élő Pachydactylus rangei nevű kis gekkó testének két oldalán, illetve a szemei körül van egy halványsárga sáv, amely 450-480 nm-es (kék) fénnyel megvilágítva 510-540 nm-es tartományban, zölden fluoreszkál. Ez a felfedezés nem pusztán azért számít kuriózumnak, mert a hüllők körében eddig leírt kékes színű fluoreszcenciától eltérően itt a kibocsájtott fény zöld, hanem azért is, mert ennek a forrása (eddigi ismereteink alapján egyedülálló módon) nem a csontszövet!
Pachydactylus rangei látható és UV fényben, felül-, oldal- és alulnézetből. Az E) képen egy tojásból frissen kikelt egyed látható, akinek testfala még olyan vékonyka, hogy azon még átüt a csontok kékes fluoreszcenciája. Ez egyébként több gekkófaj újszülöttjeinél is megfigyelhető jelenség (Prötzel et al. 2021).
A gekkók bőrének szövettani elemzése során a fluoreszkáló területeken olyan sejtekre (ún. iridofórákra) lettek figyelmesek a kutatók, amelyekben guanin kristályok halmozódtak fel. Ilyen sejteket más hüllőknél, kétéltűeknél és halaknál is találhatunk a bőr irharétegében, ott azonban csak egy ezüstös csillogást adnak az állat bőrének, esetleg a fényt megtörve és visszaverve interferencia színeket hoznak létre. A Pachydactylus rangei esetében azonban nyilvánvaló, hogy a zöldes fluoreszcencia forrása ez a sejttípus. A gekkó iridofóráinak alaposabb vizsgálata révén felismerték, hogy ezek valójában két csoportra oszthatók: egy nem fluoreszkáló, a bőr mélyebb rétegeiben található, és egy fluoreszkáló, a felhámhoz közelebb elhelyezkedő típusra. Mi a különbség e két sejttípus között, ami miatt csak az egyik képes fluoreszkálni? Nos a választ még nem ismerjük, de a cikk szerzői úgy vélik, hogy a guanin kristályok szerkezete jelentheti a kulcsot. A guanin egyébként önmagában is egy fluoreszcens molekula, de normálisan csak 170 és 290 nm-es távoli UV-vel gerjeszthető, és 350 nm körül emittál szabad szemmel még láthatatlan UV sugarakat. Elképzelhető, hogy a P.rangei zölden fluoreszkáló sejtjeinek guaninkristályaiban olyan molekuláris elrendeződés jön létre, ami kiterjedtebb π-elektron rendszer kialakulását, s ezáltal a spektrális sajátosságok megváltozását eredményezi? Talán igen. Az minden esetre biztos, hogy a bőr mélyebb rétegeiben lévő, nem fluoreszkáló iridofórák egyfajta tükörként funkcionálnak, ami a fluoreszkáló sejtek mellett elhaladó, azokkal kölcsönhatásba nem lépő fényt képes visszaverni, s ezáltal fokozni azok gerjesztő fénynek való kitettségét. Akárhogy is vesszük, ez egy ritka elegáns megoldás az anyatermészettől.
Na jó, de minek ez az egész?
A P.rangei egy éjszaka aktív sivatagi faj. Nem túl meglepő módon, a holdfényben jelen vannak olyan hullámhosszú fénykomponensek is, amelyek képesek előidézni a fent leírt zöldes fénytüneményt. Ez azonban (és itt vessünk egy pillantást a fenti képekre) nem az állat háti oldalán jelentkezik, a sötétben ólálkodó sakáloknak, vagy a magasból orvul lecsapó baglyoknak jelezve az uzsonna pontos térbeli helyzetét, hanem a test ventro-laterális részén. A mi kis gekkónk háti oldala terepmintás, felülnézetből az állat éjjel és nappal egyaránt alig észrevehető. A zöld fény a sivatag homokjából nézve, gekkó-szemszögből látható! Hogy pontosan mit üzen ez a fényjelzés, azt az etológia és a viselkedés-ökológia szakértőinek kell majd kikutatniuk, de hogy kiknek szól, az egyértelmű: a sivatag sötétjében szaladgáló fajtársaknak.
_____________
David Prötzel et al. Neon-green fluorescence in the desert gecko Pachydactylus rangei caused by iridophores. Scientific Reports (2021).
Sandra Goutte et al. Intense bone fluorescence reveals hidden patterns in pumpkin toadlets. Scientific Reports (2019).
David Prötzel et al. Widespread bone-based fluorescence in chameleons. Scientific Reports (2018).
Carlos Taboada et al. Naturally occurring fluorescence in frogs. PNAS (2017).
A napokban nagyot futott a hazai sajtóban is az Elysia chlorotica nevű tengeri csiga fantasztikus regenerációs képessége, amelynek csak a feje képes létrehozni a testet. Ugyan sokat nem tudok a sztorihoz (egyelőre) hozzáfűzni, de mivel az Elysia régi kedvencünk, azért egy rövid posztot megér a dolog.
A lenyűgöző regenerációs képesség nem az egyetlen dolog, ami miatt az Elysia híres. Ott van rögtön a kleptoplasztia, vagyis az a képesség, hogy az elfogyasztott, fotoszintetizáló algák kloroplasztiszait (színtestjeit) a bélrendszerük sejtjeivel felvegyék és tovább működtessék. Ez részben a zöld színűket is magyarázza, meg persze azt is, hogy egy fantasztikus új energiaforráshoz jutnak (kvázi “napenergiával” is működnek.)
Ehhez kapcsolódik, a másik különcségük: az algában a plasztiszok működéséhez szükség van olyan génekre ia, amit nem a sejtszervecske saját DNS-ében van jelen, hanem az alga sejtmagjában. Ezek nélkül problémás lenen a hosszadalmas fenntartásuk, de az Elysia chlorotica erre is talált megoldást. A csiga ugyanis az egyike azoknak a kevés eukarióta fajoknak, ahol bizonyítottan lezajlott horizontális gén transzfer egy másik euakrióta organizmussal és az alga legalább egy, a plasztisz fenntartásához szükséges génje átkerült a genomba.
Ezek mellett a fiatal Elysia-k fantasztikus regenerációs képessége már csak hab a tortán. Azért hangsúlyozom ezt egyébként, mert az egy évnél idősebb állatok esetében mindez már nem megfigyelhető, ott a fej levállása után nem táplálkozik és tíz nap múlva elpusztul. Maga a test akár hónapokig is életben marad (fotoszintetizál, ugye), reagál is valamennyire ingerekre, de sosem növeszti vissza a fejet.
A furcsa szokás (szakzsargonban autotómia) minden bizonnyal azért jött létre, hogy a csiga meg tudjon parazitáitól szabadulni (ezek többnyire evezőlábú rákok, a cikk szerint), hiszen azok így a régi testben maradnak és a fejből regenerálódó állat tiszta lappal indul.
Az Elysia 22 nap alatt felismerhető mértékben képes regenerálni a testét. (Mitoh és Yusa, 2021 Curr Bio)
A puhatestűek esetében egyébként nem szokatlan regeneráció jelensége: a fejlábúak esetében komoly, évszázados tudományos irodalma van a különböző csápok regenerációjának. Magyarán az alap “regenerációs eszköztár”, amire az Elysia is támaszkodhatott.
Épp ezért szokatlan, hogy a cikk megáll a jelenség puszta leírásánál. Furcsa helyzet ez, mert én magam sem szeretem azt az iskolát, ahol minden létező jelenség leírásához “fiziológiai relevanciát”, meg molekuláris adatokat követelnek a bírálók: néha egy-egy megfigyelés, részletesen leírva, önmagában is nagyon fontos. Viszont itt tényleg csak a felszínt kapirgálták meg és azért a Current Biology esetében, így 2021 magasságában, ennél több szokott kelleni a publikációhoz.
Hiszen még csak azt sem lehet mondani, hogy nem ismerjük a faj genomját és ezért nehéz mondani bármit is arról, hogy milyen gének aktiválódnak a regeneráció. Két évvel ezelőtt kijött az Elysia chlorotica genom is, így egy egyszerű transzkriptóma-elemzés most azért belefért volna. Nincs kétségem, hogy hamarosan megcsinálja azt is valaki (ha tippelhetnék, már folyamatban is van), de most még várnunk kell rá.
Mitoh S., Yusa Y. (2021) Extreme autotomy and whole-body regeneration in photosynthetic sea slugs. Curr Biol 31(5): R233-R234. doi: 10.1016/j.cub.2021.01.014.
Cai, H., Li, Q., Fang, X. et al. (2019) A draft genome assembly of the solar-powered sea slug Elysia chlorotica. Sci Data6, 190022. https://doi.org/10.1038/sdata.2019.22
Az Amerikai Egyesült Államok leggyakoribb ízeltlábú köztigazda által terjesztett betegsége a Lyme-kór, évente háromszázezer embert fertőz meg. A betegséget kullancsok terjesztik, Amerikában főleg a feketelábú kullancs (Ixodes scapularis). Azonban az emberi megbetegedések rejtélyes eloszlást mutatnak, mivel eddig sehogyan sem sikerült őket fedésbe hozni a kullancsok eloszlásával. Bár a köztigazdaként működő kullancsfaj az USA északkeleti és északi részétől egészen a Mexikói-öbölig elterjedt, Texasban és Floridában is, ám a Lyme-kór emberi áldozatai az északi területeken sűrűsödnek, a déli államokban a betegség kifejezetten ritka. Sokáig nem találtak meggyőző választ a kérdésre, hogy miért különbözik a Lyme-kór eloszlása északon és délen, ha a feketelábú kullancs egyformán elterjedt mindkét területen?








.jpg){kind=link}
{kind=link}